ЭКОЛОГИЯ ЕЛОВЫХ ЛЕСОВ
Введение
Изучение экологии лесов, и в частности еловых, имеет большое значение для продвижения их на север, в ныне безлесные места, для разработки мероприятий по восстановлению еловых лесов, повышению их продуктивности. Важность экологических исследований бесспорна. Имеется много примеров отрицательных результатов деятельности человека, проводившего хозяйственную деятельность, не зная или не учитывая экологию леса.
Распространение еловых лесов, их фитоценотическая устойчивость, продуктивность и качество древесины связаны с определенными экологическими условиями. Обширный ареал еловых лесов приводит к их большому диапазону.
Так, среднегодовые температуры воздуха изменяются от -2,9 до +4°С.
Средняя температура самого теплого месяца (июля) от 10 до +20°С. Про-
должительность вегетационного периода 110-175 дней. Сумма температур
выше 5°С изменяется от 700 до .2230°, выше 10°С-от 500 до 1850°. Коли-
чество осадков 400-850 мм. Среднемесячная относительная влажность воз-
духа в 13 ч от 53 до 88%. Число часов солнечного сияния в среднем изме-
няется от 1455 в предтундровой до 1585 в среднетаежной подзоне.
При всем разнообразии показателей можно выделить общее:
Еловые леса приурочены к зоне прохладного и влажного климата, гидротермический коэффициент (по Селянинову) изменяется от 0,57 до 0,38. Показатель увлажнения (по Шашко) высокий и составляет по подзонам тайги 0,7-0,6. Радиационный индекс сухости (по Будыко, 1956) колеблется от 0,85 до 1. Это зона четко выраженных времен года с холодной снежной зимой и умеренно теплым, с достаточным количеством осадков летом. Количество осадков превышает потенциальную возможность их
испарения. Влажность воздуха достаточно высокая.
Еловые леса произрастают преимущественно на подзолистых почвах, умеренно теплых и достаточно увлажненных в течение
всего вегетационного периода.
Еловые леса заметно влияют на климат и почву, трансформируют их в благоприятном для себя направлении как с точки зрения обеспечения энергии роста. Так и с точки зрения конкурентных отношений с другими видами растений, за выживаемость ели как вида.
Ниже будут рассмотрены некоторые элементы микроклимата (фитоклимата) еловых лесов, их связь с типами лесных
биогеоценозов, с ‚типами леса, трансформация элементов климата под влиянием еловых лесов, а также взаимоотношения
древесной растительности и почвы. Основой всех процессов яв-ляется солнечная энергия.
Радиационный режим
Лучистая энергия солнца-основа всех жизненных процессов на земле.
В среднем к поверхности атмосферы ежеминутно поступает 7,88 дж на 1 см. Поверхности же живого покрова земли достигает значительно меньше энергии. Она частично поглощается и рассеивается атмосферой.
Таежная зона характеризуется такими величинами годовой суммарной радиации:
от 293 до 356-377 кДж/см2-год (Данилова, 1962). Причем за май-сентябрь поступает 0,76-О,72%
общей годовой радиации. А за один июнь поступает 0,18-0,2 годового количества тепла. В поступлении тепла в таежной зоне существует определенная сезонность. Величина эффективного излучения при безоблачном небе (максимально возможное излучение) колеблется от 142 до 168 кДж/смг-год‚ что составляет 40-45% годичного прихода тепла. За вегетационный период отражается 43-45% годового количества отраженного тепла.
Еловые древостои тайги в значительной степени изменяют и перераспределяют поток солнечной энергии. Так, псд полог среднетаежных ельников-чернйчников нормальной сомкнутости (полнота 0,7-0,9, сомкнутость крон ~0,7) поступает лишь 9-
15% суммарной радиации в среднем за сезон, под полог северотаежных таких же ельников-черничников-7-130/0. ПОД полог травяно-болотных ельников поступает на 3-40/0 больше суммарной радиации. Это объясняется строением крон ели
в этих условиях. Причина меньшей интенсивности потока солнечной радиации, поступающего под полог северотаежных ель-
ников, меньшая высота солнца B течение всего вегетационного периода. Остальная часть солнечной энергии поглощается и отражается кронами елового древостоя. Средняя величина альбедо хвойных лесов равна 14%. На процессы жизнедеятельности ели, на прогревание крон, стволов деревьев, а также
воздуха тратится около 70% приходящей солнечной радиации.
Из поступающей под полог еловых лесов энергии солнца
около 20-30% отражается от живого напочвенного покрова и почвы. Для жизнедеятельности нижних ярусов леса и на прогревание почвы расходуется 7-100/0 полной суммарной солнечной радиации.
Многолетние наблюдения за интенсивностью солнечной радиации показали, что зависимость интенсивности суммарной радиации под пологом ельников от интенсивности суммарной радиации на открытом месте выражается уравнением параболы 111 степени. Коэффициент корреляции колеблется от 0,3 до 0,7
в зависимости от времени суток, состояния облачности и т. п.
Эта зависимость действительна для периода, когда лиственные породы, входящие в состав древостоя, находятся в облиственном состоянии. Весной, а также осенью, с началом листо-пада интенсивность суммарной радиации пол пологом ельников несколько возрастает. Интенсивность солнечной радиации за-
висит от высоты солнца и продолжительности дня, поэтому наибольшие значения ее как на открытом месте, так и в ельниках отмечаются в июне, а в августе они значительно уменьшаются.
максимальное значение суммарной радиации на ОТКРЫТОМ месте за время
наблюдений составляло 5,28 Дж/см2-мин (ясные дни), минимальное-
0,4 Дж/см2- мин (пасмурные дни), что составляло 8,1 % величины в ясные дни.
Очень важный показатель-количество прямой солнечной
радиации, поступающей пол полог леса. Отсутствие прямых
солнечных лучей тормозит прохождение фаз растениями нижних
ярусов леса, они часто не цветут. Удельный вес прямой радиации B суммарной, поступление ее под полог древостоя зависят
от характера погоды. В ясные дни основной составляющей суммарной радиации является прямая радиация (табл. 4‘), B nacмурные-возрастает доля рассеянной, которая вследствие отсутствия определенной направленности легче проникает в глубь леса. По сравнению со средними данными напряжение суммарной радиации в ясные и пасмурные дни значительно изме-
няется:
B ясные дни оно приблизительно в 1,5-2 раза больше
среднего, в пасмурные-в 1,5-2 pasa меньше. Интенсивность
суммарной ипрямой радиации снижается от июня к сентябрю.
Так, в июне она составляла на открытом месте 100%, в июле 86%, в августе - 70%.
B пасмурные дни, которые преобладаютвтаежной зоне, доля
прямой радиации на открытом месте невелика, под пологом
ельников часто отсутствует. В такие дни уменьшается разница между интенсивностью радиации на открытом местеи
B ельниках, где она может достигать 30-35% интенсивности
суммарной радиации на открытом месте. Под пологом ельников
большую часть вегетационного периода господствует рассеянная радиация, бедная физиологически активными лучами.
Расчеты количества тепла, приходящего за один день ясной погоды, показали, что под полог ельников-черничников среднесуточные величины суммарной радиации при ясной и пасмурной погоде отличаются: в ясные дни количество тепла
под пологом ельника-черннчннка свежего составляет 6-10% количества тепла на открытом месте. В пасмурную погоду под
пологом ельника-черничника свежего суточные суммы суммарной радиации составляют около 20%. B ясную погоду B 13 ч отмечается выраженный максимум изменения интенсивности суммарной радиации, в пасмурную погоду суточный диапазон выражен слабее.
В таежной зоне под полог ельников проникает 7-150/0 преимущественно рассеянной суммарной радиации.
Температура воздуха
Температурный режим ельников непосредственно связан с поступлением поц полог еловых лесов солнечной радиации.
Температура воздуха в таежной зоне обусловливается приходом теплых воздушных масс из Атлантики и Средиземноморья.
Одновременно наблюдаются и более локальные процессы в лесных биогеоценозах.
Источник нагрева воздуха в лесу в течение дня - нагретые
лучами солнца кроны деревьев, стволов и участки почвы, на которые попадают прямые солнечные лучи. Площадь и характер нагрева зависят от вертикальной и горизонтальной сомкнутости полога. Показатель этой сомкнутости, а также площади
почвы, нагреваемой лучами солнца, характеризуется абрисом
солнечных бликов, площадь которых не превышает 10%. Благодаря этому температура воздуха под пологом ельников в течение вегетационного периода всегда ниже, чем на открытом ме-сте (табл. 7). Наибольшая разница наблюдается между температурами в приповерхностном слое воздуха на открытом ме-
сте и в ельнике травяно-сфагновом, где она достигает 2-3°С
(табл. 8). Это объясняется большей прогреваемостью воздуха
на обезлесенных участках за счет тепла от поверхности почвы.
В лесу воздух нагревается от стволов, ветвей, заходит с без-
лесных площадей. К поверхности почвы тепла поступает мало и большая часть его из приповерхностного слоя воздуха расхо-
дуется на прогревание почвы. В результате у поверхности почвы
воздух обычно хололнее, чем на высоте 1,5 M, т. е. наблюдается
инверсия температур. Лишь осенью, с началом интенсивйого
охлаждения воздуха и почвы, и под пологом ельников воздух
бывает теплее y поверхности почвы. Такое изменение темпера-
тур наблюдается во всех подзонах тайги.
B годы с более холодным вегетационным периодом умень-
шается разница температур воздуха ПО ВЫСОТЗМ, СВОДИТСЯ K MH-
HHMyMy И разница между ТИПЗМИ леса.
Наиболее резко различаются температуры воздуха на откры-
TOM месте И В ельнике В ЯСНУЮ солнечную ПОГОДУ (антицикло-
нальный тип ПОГОДЫ), особенно утром, когда разница в темпе-
ратурах достигает 5-6°. Днем и вечером различия уменьша-
ются, B пасмурную погоду (циклональный тип погоды) сглаживаются.
Статистическая обработка данных 5-летних наблюдений показала тес-
ную связь температур воздуха в лесу и на открытом месте (коэффициент
корреляции равен 0,857-0,966).
Уравнения связей .среднедневных температур между открытым местом
х и ельником травяно-сфагновым y (средняя подзона тайги) имеют следую-
щий вид:
в ясную погоду
y=-3,04+0,992x } 7 ч на 0,0 M
y=-l,73+0,924x В на 1,5 M
y=-1,68+0,95ox } в l3 ч на 0,0 M
y=0,574+0,957x ‚ на 1,5 M
y=0,84+0,873x } в 19 ч на 0,0 M
y=0,91+0,875x на 1,5 м
в пасмурную погоду
у=0‚36+0‚887х } 7 ч на 0,0 м
y=0,41+0,905x В. на 1,5 M
y=0,56+0,896x } в 13 ч на 0,0 M
y=0,757+0,938x Ha 1,5 M
y=0,103+0,941x } в 19 ч на 0,0 M
y=0,77+0,90ox на 1,5 M
Уравнения связей среднедневных температур воздуха между открытым
местом x- и ельником-черничником свежим у1 имеют следующий вид:
в ясную погоду
У1=-0‚82+0‚948х } в 7 ч на 0,0 M
y1=0,13+0,911x на 1,5 M
y;=-l,63+0,988x на 0,0 м
у",=0‚20+0‚986х } в ‘3 “ на 1,5 M
y1=0,04+0,956x на 0,0 M
yz=0,70+0,918x } В 19 Ч на 1,5 M
в пасмурную погоду
У1=0‚36+0‚934х } в 7 ч на 0,0 м
У1=0‚273+0‚947х на 1,5 M
У1=-0‚06+0‚907х} I3 на 0,0 M
y1=0,513+0,926x B Ч на 1,5 м
yl=0,424+0,928x } в 19 ч на 0,0 м
уі=0‚935+0‚896х
Температурный режим ельников зависит и от их типа, отражающего весь комплекс лесорастительных условий. В заболо-
ченных типах леса (ельник травяно-сфагновый) весной воздух
немного холоднее, чем в суходольных типах леса (ельник-черничннк свежий). Летом такая закономерность отмечается лишь
для приповерхностных слоев воздуха (0-10 см), на высоте
l-l,5 м_ четкой разницы не наблюдается (табл. 9).
Осенью, с началом выхолаживания, воздух в ельнике тра-
вяно-сфагновом становится холоднее, чем в черничнике свежем.
Разница в температурах воздуха между открытым местом и в ельнИках, особенно в травяно-сфагновых, достигает наибольшей величины
в начале вегетационного периода. K осени эта разница уменьшается. На высоте 1-l,5 M разница температур воздуха на от-
крытом месте и в ельниках значительно меньше, чем в приповерхностном слое воздуха. В конце вегетационного периода она
увеличивается. Существенно изменяются температуры воздуха
в разное время суток в различных типах леса. В утренние часы
(наблюдения в 7 ч) в приповерхностных слоях воздуха и на высоте 1,5 M температура в черничниках характеризуется быстрым
и сильным увеличением в периоды потепления и сильным снижением B холодные периоды.
B ельниках-черничниках наибольшие максимальные темпера-
туры отмечаются на высоте 1 м. У поверхности почвы они имеют
меньшую величину. Разница в среднем за месяц составляет
1.‚3-1‚8°С.
В ельниках травяно-сфагновых наибольшее значение максимальных температур наблюдается на высоте 0,5-1 м, разница
составляет 2,5-3°С. На открытом месте максимальные температуры имеют наибольшее значение у поверхности почвы.
Наиболее сильное охлаждение в ночное время наблюдается
B ельнике травяно-сфагновом (рис. 8). Эта зависимость сохраняется для высоты 1-1‚5 M и для приповерхностного слоя
воздуха.
На открытом месте самые низкие температуры наблюдаются на
высоте 0,25 M, т. е. на поверхности травянистой растительности.
В ельнике травяно-сфагновом наиболее низкие минимальные
температуры отмечаются на высоте 1 м.
Редко бывающие, но захватывающие все типы
леса адвективные заморозки вызываются вторжением холодных
масс воздуха. В лесу они слабее, чем на открытом месте. Наи-
более сильные заморозки, преимущественно адвективные, от-
мечаются на открытом месте, где температура воздуха y по-
верхности почвы Может опускаться до-8-1О°С. B- ельниках
тр"авяно-сфагновых заморозки значительно слабее-До -3,З°С.
В ельниках-черничниках свежих за 5-летний период наблюде-
ний минимальные температуры приповерхностного слоя воздуха
не опускались ниже 2-2‚5°.

14. Наибольшая сила заморозков на поверхности почвы
(средняя подзона тайги).
На открытом месте адвективный заморозок наиболее силен на поверхности травяного покрова (0,25 м). В ельнике травяно-сфагновом, расположенном в пониженном месте, заморозок
сильнее, чем в черничнике, наиболее низкая температура отмечена на высоте 1 м. В приповерхностном слое воздуха, граничащем с водой, имеющей большую теплоемкость, значительно теплее. В ельнике-черничнике по всему вертикальному профилю величина отрицательных температур примерно одинакова.
Влажность воздуха
Влажность воздуха имеет существенное значение для жизии
древесных растений. Признано, что ель может произрастать
лишь в местностях с высокой влажностью воздуха, сухость воздуха губительна для нее. Влажность воздуха связана с тем-
пературой и движением воздуха, a также другими элементама климата.
Для характеристики влажности воздуха обычно используются
такие показатели: относительная влажность и дефицит влажности воздуха. Относительная влажность воздуха показывает степень насыщенности воздуха водяными парами.
Дефицит влажности воздуха-разность между максимальной упругостью водяного пара, насыщающего пространство при данной температуре, и действительной упругостью.
От дефицита влажности воздуха зависит испарение расте-
ния. При благоприятной влажности воздуха транспирациониый
аппарат растения работает нормально: недостаток влажности
вызывает угнетенное состояние растения. Фотосинтез наиболее
успешно протекает при относительной влажности воздуха, до-
стигающей 60-70%. Относительная влажность воздуха во мно-
гом зависит от состояния погоды: в период устойчивой и ясной
погоды суточные колебания ее выражены резче. При этом амплитуда колебаний влажности в ночные и дневные часы значительно больше, чем при неустойчивой погоде. В пасмурную погоду амплитуда колебаний незначительна. Днем значения относительной влажности воздуха минимальные, ночью они повы-
шаются, нередко достигая 100%.
Наибольшая разница между относительной влажностью под
пологом ельников и на открытом месте отмечается в предутренние часы, при этом влажность воздуха в ельниках всегда выше.
Между 6-8 ч показатели влажности воздуха выравниваются.

B дневное время влажность воздуха на открытом месте обычно
ниже, чем под пологом ельников. Минимальное значение относительной влажности отмечается между 13-16 ч. Время наступления суточного минимума относительной влажности воздуха совпадает со временем суточного максимума температуры. Наименьшая влажность в течение всего вегетационного периода наблюдается на обезлесенных участках как у поверхности почвы, так и на высоте 1,5 M. Наибольшая влажность в ельнике травяно-сфагновом. Разница влажности воздуха на открытом
месте и B ельнике травяно-сфагновом составляет 10-15%.
В 7 ч влажность воздуха“ в ельнике травяно-сфагновом иногда
достигает 96-99%. 3a весь сезон среднедневная относительная
влажность приземного слоя воздуха B травяно-сфагновом ельнике на 3-10% выше, чем в ельнике-черничнике. На высоте 1,5 м разница колеблется от 1 до 4% (табл. 16).
В течение вегетационного периода наименьшие значения относительной влажности воздуха наблюдаются в мае (50-85%у поверхности почвы и 47-70% на высоте 1,5 м). Затем значения влажности воздуха постепенно возрастают и достигают
в августе 85-92% y поверхности почвы и 80-85% на высоте
1,5 м (табл. 17). Такой ход изменения относительной влажности воздуха свидетельствует о том, что в мае-июне создаются
наиболее благоприятные условия для испарения влаги и транспирации. К осени эти процессы ослабляются. В холодный веге-
тационный период с количеством осадков выше средней нормы
значения влажности высокие весь период, разница между типами леса, а также лесом и открытым местом минимальная.

Разница относительной влажности воздуха у поверхности
почвы и на высоте 1,5 м составляет: на открытом месте 7-9%‚
в ельнике-черничнике-6-7%, B ельнике травяно-сфагновом-
9%. В 7 ч в июне-июле на открытом месте она равна 4%,
в черничнике травяно-сфагновом - 6%. В 13 и 19 ч разница увеличивается до 8% на открытом месте и в черничниках и до 11% в травяно-сфагновых. Понижение в отдельные периоды влажности воздуха в ельниках-черничниках свежих по срав-
нению с влажностью на открытом месте объясняется выпадением осадков малой интенсивности. На открытом месте они вызывают повышение влажности воздуха, под полог ельннков они не проникают и не влияют на влажность воздуха.
Дефицит влажности наиболее проявляется на открытом месте, где он может достигать 1000-1300 Н/м2. Под пологом ельников он обычно не превышает 1000-1100 Н/м2. В меньшей
степени дефицит влажности проявляется B ельнике травяносфагновом, в большей - в черничнике свежем (табл. 18). В течение суток дефицит влажности имеет наибольшие значения днем, наименьшие утром. И в ельниках и на открытом месте
дефицит влажности у поверхности почвы меньше, чем на высоте 1,5 M.

Под пологом ельников влажность воздуха всегда более вы-
сокая, Т. е. создаются условия для пониженной транспирации
подростом ели, ЧТО способствует повышению его ЖИЗНЕСПОСОБНОСТИ В конкуренции с материнским пологом.
Температура почвы
Температура Почвы имеет важное значение в жизни биогео-
ценозов таежной зоны. Температура почвы определяет возмож-
НОСТЬ прорастания семян И появление ВСХОДОВ лесных РАСТЕНИЙ.
OT нее в значительной степени зависят строение корневой си-
стемы, характер и интенсивность роста корней, возможность
иинтенсивность потребления растением из почвы подвижных
форм элементов питания, особенно азота.
Температура почвы определяет интенсивность деятельности
почвенных животных и микроорганизмов, что B свою очередь
обеспечивает успешность переработки органического вещества
отпада, перевола элементов питания в форму, доступную для
усвоения растениями. В первую очередь это касается процессов аммонификации и нитрификацни.
В температуре почвы, как и в других элементах климата,
обнаруживается тесная связь с географическими условиями,
выражаемыми через лесорастительные педзоны. В ельниках предтундровой, северной и среднетаежной подзон отмечена пдентичность изменений температуры почвы в течение вегетационного периода (рис. 11). Наименьшие значения температур почвы наблюдаются B предтундровых ельниках, по мере продвижения к югу температура почвы повышается.

Таежные еловые леса B значительной степени влияют на температурный режим почвы. Почвы под еловыми лесами обычно
всегда холоднее, чем пед сосняками и мелколиственными ле-
сами. Интенсивное прогревание почвы весной в ельниках и на
вырубках начинается неодновременно из-за более позднего
схода снега и более позднего оттаивания почвы B лесу. B это время различия B температуре почы на открытом месте
значительно возрастают и в июне достигают максимальной величины. Средняя за июнь температура почвы в северотаежном
ельнике на глубине от 5 до 40 см на 7-8°С‚ а на 80-120 см
на 5-6°С ниже по сравнению с, температурой на открытом месте. В средней подзоне тайги различия температур почвы на
глубине 5-20 см на открытом месте и в ельнике B Mae составляют 3-5°С, B июне 4-5°C. B июле и августе различия
в температуре почв уменьшаются по сравнению с температурами
B июне и составляют 3-5°С B ельнике северной тайги и 2-4°С
в ельниках средней тайги.
Осенью вследствие более быстрого охлаждения почвы на открытом месте по сравнению с лесом различия B температуре почвы на этих участках снова уменьшаются. Зимой (январь-›-март) на глубине 20 см температура почвы B ельниках ниже
на 0,7-1‚8°С (в среднем за январь 2,2°С). На глубине 1,2 M
разница температур достигает 1-1,2°С.
Самые холодные почвы в это
время обычно в травяно-сфагновом ельнике. Это объясняется
тем, что веда, которая в это время в травяно-сфагновом ель-
нике стоит на поверхности, имеет высокую теплоемкость и по-
этому прогревается медленнее. Также медленнее оттаивает и
почва.
На открытом месте верхние 20 см почвы прогреваются 15-
25 мая до 6-7°С. Под пологом ельников-черничников этот пери0д обычно наступает 20-25 июня, в заболоченных травяно-сфагновых ельниках в конце июня - начале июля, т. е. под по-логом среднетаежных ельников-черничников прогревание почвы
запаздывает почти на месяц, а псд пологом травяно-сфагновых
на 30-40 дней.
Верхний 20-сантиметровый слой почвы, где сосредоточена
основная масса корней древесных пород, прогревается в северной подзоне тайги до температуры 6-7°С на открытом месте
в последней пятидневке мая, B березняке- 11-15 июня, в ельниках-в третьей декаде июня. Таким образом, прогревание
почвЫ до температуры, необходимой для начала активного роста корней, в северотаежных ельниках запаздывает на месяц
по сравнению с открытым местом.
Неодноролность сомкнутости древесного яруса и хорошо
развитый микрорельеф в ельниках обусловливают сильные ко-
лебания температур почвы по площади. Например, в ельниках-черничниках колебания составляют 6-8°С, B травяно-сфагно-
вьтх-5-9°С.
«Температурные карты», составляемые несколько раз за ве-
гетационный период, показывают большую пестроту температур
почвы в лесу. Наиболее высокие температуры почвы обычно
наблюдаются на микроповышениях, в местах больших просветов
в пологе крон. Прогалины в словом лесу способствуют прогре-
ванию почв. На прогалине, на расстоянии 1-2 M OT стены леса
на север, изменение температур почвы не обнаруживается. Но
на расстоянии около 10 м от края леса температуры почвы за-
метно увеличиваются. В ельнике-черничнике почва прогревается
до 6°С на глубине 30 см, в прогалине-до глубины 50 см.
В микропонижениях обычные низкие температуры держатся
ДОЛГО.
В южной тайге принципиальный ход температур такой же (Орлов,
1953). Наблюдаются лишь сдвиг дат начала прогревания почвы и различия
в абсолютных значениях температур.
Сезонный ход температур воздуха и почвы в ельниках пока-
зывает, что в таежной зоне, особенно в ее северной части,
почва прогревается позднее воздуха. В отцельные ГОДЫ эта
разница достигает 1 месяца, привоця к тому, что при активных
ростовых процессах В кроне корневая система. функционирует
не полностью.
Промерзание почв.
В таежной зоне обычно наблюдается
ежеголное зимнее промерзание ПОЧВ. минеральные ПОЧВЫ неза-
болоченных ТИПОВ леса промерзают В зимнее время на значи-
тельную глубину. Глубина промерзания B значительной степени
связана С ПОГОДНЫМИ УСЛОВИЯМИ осени, С ГЛУбИНОЙ снежного
покрова, МОРОЗНОСТЬЮ ЗИМЫ, влажностью ПОЧВЫ И- Т. П.
Так, в 1966-1967 гг., когда была холодная осень и малоснежная зима,
почва промерзла в ельниках (средняя подзона тайги) до глубины 100 CM
B первую декаду февраля. В 1968-1969 гг. снег выпал рано, сразу же после
начала промерзания почвы, и достиг большой мощности (57-67 см). Почва
в этот год промерзала медленно, достигнув в конце января глубины 45-
46 см в ельниках.
В 1969 г. промерзание почвы началось в конце октября одновременно
с выпадением снега и достигло к началу ноября 8-12 см B ельнике и 2 CM
на открытом месте. Повышение температуры воздуха в первой половине
ноября вызвало стаивание снега и уменьшение мерзлого слоя почвы до
6-7 см в ельнике-черничнике. На открытом месте почва оттаяла полностью.
Bo второй половине декабря вновь началось быстрое промерзание почвы.
Наиболее интенсивно шло оно в ельнике, где почва промерзала на 3-5 CM
B сутки, на открытом месте-около 1 см в сутки. Аналогичные данные по-
лучены ранее А. А. Молчановым (1952).
Максимальное промерзание наблюдается В марте-апреле.
В ельниках почва промерзает на глубину 120 см, на открытом
месте-до 48-50 CM. Глубина промерзания значительно ко-
леблется по годам - от 20 до 120 см.

Глубина промерзания почв зависит от типа леса, возраста
древостоя (Сахаров, 1938, 1948; Молчанов, 1961). В заболоченных типах лес‘а глубина промерзания меньше. Между глубиной
промерзания B различных типах заболоченных лесов устойчивых различий нет (Изотов, 1968). Это объясняется однородными ус-
ловиями промерзания: близкими условиями увлажнения, физи-
ческими свойствами верхних горизонтов почвы и в какой-то мере
незначительной разницей в мощности снежного покрова. Так,
по данным В. Ф. Изотова (1967), мощность снегового покрова
в заболоченных типах леса различалась по годам на 5-10 см.
Продолжительность периода от начала промерзания до не-
риода наибольшей глубины промерзания в ельниках-чернични-
ках колеблется от 120 до 170 дней, интенсивность промерзания
в зависимости от температурных условий зимы-от 0,15 до
1 см в сутки. На открытом месте перИОД промерзания почвы
короче-60-160 дней, интенсивность промерзания-0,1-
0,4 см в сутки.
В средней подзоне тайги почва оттаивает обычно как сверху
от талых вед и солнечных лучей, так и снизу благодаря при-
току тепла от нижележащих талых слоев почвы. В отдельные
годы (1967) оттаивание начинается сверху, иногда (1968)
снизу. В ельниках-черничниках северной подзоны тайги почва
начинает оттаивать обычно снизу. Оттаивание начинается в ап-
реле, в отдельные годы-в последних числах марта. Сверху
почва начинает таять во второй половине мая, иногда-в на-
чале мая. Обычно это совпадает с концом периода снеготаяния.
Полностью почва оттаивает в ельниках-черничниках в середине
июня. Период оттаивания почвы продолжается 60-80 дней.
В ельниках травяно-сфагновых почва оттаивает на несколько
дней позднее. На открытом месте почва оттаивает вследствие
поступления тепла из нижних горизонтов почвы. Начинается
этот процесс в конце марта-начале апреля, заканчивается
обычно к концу снеготаяния, т. е. когда псд пологом ельников
начинается уменьшение слоя мерзлоты сверху. Продолжитель-
ность периода оттаивания почвы - 35-60 дней.
В первые дни почва оттаивает на всех участках медленно
(1-2 см за пятидневку). Увеличение среднесуточных темпера-
тур воздуха в начале мая способствует возрастанию скорости
оттаивания снизу до 1-3 см и больше в сутки. Разница в вели-
чине промерзания почвы на открытом месте и в ельниках раз-
личных типов увеличивается в годы с более глубоким промер-
занием.
Осадки
Влага в жизни растений играет огромную роль. ВОДа попа-
дает в биогеоценоз в виде атмосферных осадков, в меньшей сте-
пени -за счет притока почвенных вод. Проникновение осадков
в лесу зависит от типа леса, возраста, полноты и состава дре-
востоя. Не все осадки в лесу попадают на почву, часть их за-
держивается кронами. Кроны еловых насаждений задерживают
осадков больше, чем кроны березовых или сосновых.
Большое значение в снабжении почвы водой имеют твердые
зимние осадки. Европейская тайга относится к зоне, где тол-
шина снежного покрова в среднем составляет 50-70 см (Рих-
тер, 1945). Мощность снежного покрова в еловых лесах колеб-
лется по годам от 50 до 80-85 см. В малоснежные зимы под
полог еловых лесов поступает 60-70% количества снега на
открытых местах (табл. 22). B многоснежные зимы под полог
ельников поступает 80-85 % снега.

Под полог травяно-сфагновых ельников, кроны деревьев
в которых более узкие, снега обычно поступает на 5-6%
больше. Наименьшие колебания мощности снега по площади
отмечаются на открытом месте (от 87 до 109 см, о=5‚5), наи-
большие-в заболоченных ельниках вследствие сильно выра-
женного микрорельефа (от 57 до 98 см, о=13‚3). В ельнике-чер-
ничнике свежем с относительной выравненностью рельефа эти
колебания составляют от 65 до 91 см (Or-6,3).
Плотность снега на открытом месте и в ельниках 0,20-
0,23 r/CM3.
Снег, являющийся источником ‚влаги для растений, благо-
даря своей рыхлости (уд. вес 0,1-0‚25 г/смз) и значительной
мощности служит хорошей защитой от морозов зимующих юных
древесных растений и представителей живого напочвенного
покрова, а также зимующих в подстилке животных. Коэффи-
циент теплопроводности снега очень мал-*13‚1- 36,6 1°С
(сутки/см) при плотности снега 0,5-0,25 r/CM3. Благодаря этому
разница температур воздуха и на поверхности почвы состав-
ляет при мощности снега 20 см - 10-15°С. При мощности снега
40 см и более в течение зимы на почве обеспечивается постоян-
ная температура - не ниже -10°С.

B лесу снег тает медленнее, вода задерживается на месте
вследствие выраженного микрорельефа. В результате в лесу
задерживается значительно больше талых вод, чем на откры-
тых пространствах. Интенсивность снеготаяния во многом за-
висит и от географического положения: продолжительность сне-
готаяния на безлесных участках B северной псдзоне тайги составляет 20-24 дня, в южной около 20 дней. Продолжитель-
ность таяния под пологом ельников примерно в 1,5 раза больше
продолжительности снеготаяния на полянах.
Интенсивность снеготаяния в дружные весны составляет за сутки в ело-
вых лесах 5 мм, в сосновых -"7 мм, в лиственных- 10 MM, на лесных поля-
нах 12-16 мм (Рутковский, 1948).
Больше подовины влаги поступает в почву в виде жндких
осадков. Лес и здесь влияет на количество осадков, достигаю-
щих почвы (рис. 15). Ельник в среднем за вегетационный пе-
риод задерживает в средней подзоне тайги 32-46% осадков,
в северной подзоне тайги 30-40% осадков (рис. 16). Количе-
ство осадков, попадающих B почву в ельниках, зависит от продолжительности и интенсивности дождя. Во время высоко ин-
тенсивных дождей под полог ельников поступает 70-85% осад-
ков. Моросящие низкоинтенсивные дожди сильно задержи-
ваются кронами деревьев и non полог их поступает 10-
20%.
При дождях интенсивностью 1-2 мм под пологом древостоев может
быть учтено больше осадков, чем на открытом месте. Это явление объяс-
няется конденсацией ведяных паров, которая больше, чем выше сомкну-
ТОСТЬ крон. степень проникновения ОСЗДКОВ ПОД ПОЛОГ древостоев связана
с сомкнутостью крон: чем более они сомкнуты, тем меньше осадков посту-
пает К поверхности ПОЧВЫ. В светло-
ХВОЙНЫХ лесах эта ЗЗВИСИМОСТЬ пря-
молинейна.
Ветер
Роль движения воздуха, или ветра, в жизни леса трудно пе-
реоценить. Ветер приносит B лессобезлесенных участков тепло,
благодаря которому происх0дит таяние снега и прогревание
почвы весной. Ветер способствует более интенсивному обмену
воздуха, увеличению транспирации, замене обедненного угле-
кислотой воздуха воздухом богатым содержанием Co2. Ветер
усиливает фотосинтетическую деятельность деревьев, содейст-
вует испарению c поверхности почвы в лесу, изменяет влаж-
ность и температуру воздуха, перенося семена растений, спо-
собствует их расселению, а являясь единственным переносчиком
пыльцы хвойных древесных растений, обеспечивает их семен-
ное размножение. Однако ветер большой скорости увеличивает
транспирацию, уменьшает фотосинтез, что приводит к сниже-
нию прироста деревьев в высоту.
Ветер на севере таежной зоны заметно влияет и на форми-
рование крон у деревьев. На границах с открытым местом, осо-
бенно по побережью северных морей, у деревьев формируется
флагообразная крона. На побережье“ Белого моря крона у де-
ревьев вытянута в южную, юго-восточную сторону. Наиболее
сильное влияние на отмирание ветвей кроны оказывают север-
ные и северо-западные ХОЛОДные ветры. В ряде случаев наряду
с образованием флагообразной кроны происходит и усыхание
вершин, что наиболее выражено у ели.
В условиях Архангельской области B течение вегетационного периода
преобладают ветры западного направления (З, СЗ, ЮЗ). В мае также часто
повторяются северные и северо-восточные ветры. В июне-ветры южных на-
правлений, В ЗВГУС"Ге- ВОСТОЧНЫХ И ЮГО-ВОСТОЧНЫХ.
Наибольшая сила ветра наблюдается днем, к вечеру ветер стихает. По
наблюдениям в 7 ч среднемесячная скорость ветра колеблется от 0,77 до
1,9 м/с, в 13 ч-от 1,95 до 3,1, B 19 ч-0,47-2,7 м/с.
Лес, являясь сильным препятствием на пути ветра, значи-
тельно ослабляет его силу. Скорость ветра в ельниках-чернич-
никах составляет 12-20% скорости на открытом месте (5-
8 м/с). Скорость ветра в лесу изменяется в зависимости от вы-
соты над поверхностью почвы. Самые низкие скорости ветра от-
мечаются над поверхностью почвы, с увеличением Высоты они
возрастают.
Велика роль ветра как сильного агента межбиогеоценозного
обмена веществ. Весной ветер переносит большие количества
цветочной пыльцы из одного биогеоценоза в другой. Так, масса
пыльцы сосны составляет 11-65 кг/га сосновых насаждений
ежегодно (Журавлева, 1974). Осенью ветер переносит усохшие
листья, мелкие веточки, семена и т. п. Одновременно перено-
сятся большие количества пыли, газов.
B современный перн0д атмосферная пыль, выпадающая на
поверхность почвы, служит одним из важных факторов почво-
образования. Количество выпадающих ‘зольных веществ соиз-
меримо с количеством их, требующихся на построение годич-
ного прироста фитомассы в некоторых биогеоценозах.
Движение воздушных масс в лесу-основная причина рас-
пространения паразитных и сапрофитных грибов. В заповедном
‚перестойном лесу в 1 м3 воздуха содержится до 1995 спор гри-
бов, а в пройденном рубками ухода лесу - около 500-700 спор.
Эта масса спор разносится ветром из одного участка леса в дру-
гой (Черемисинов, 1974).
Велика роль ветра и в увеличении псд пологом леса пло-
щади, непосредственно получающей прямую солнечную радиа-
цию. Ветер способствует образованию движущихся солнечных
бликов благодаря раскачиванию крон деревьев, что почти вдвое
увеличивает площадь, на которую падают прямые солненые
лучи. Следовательно, ветер является одним из факторов, суще-
ственно определяющих радиационный режим леса.
Почва
Почвенные условия в жизни леса имеют не менее важное
значение, чем климатические факторы. В определенных клима-
тических условиях от свойств почвы зависят породный состав
леса, быстрота роста и продуктивность древостоев, качество
древесины и т. п. В свою очередь почва, как указывал осново-
положник генетического почвоведенйя В. B. Докучаев (1949),
представляет собой тело, образовавшееся в результате сово-купной деятельности и влияния материнской породы, расти-
тельных и животных организмов, климата, возраста страны
и рельефа местности. Это синдетельствует о сложных взаимо-
отношениях между растительностью и ночной.
Зона тайги должна совпадать с почвенной зоной, особенно
если последнюю понимать как почвенно-климатическую зону
(Соколов‚_ 1968). Зональные почвы для тайги-подзолистые.
По механическому составу подзолистые почвы представлены
всеми разностями, начиная от песков и кончая глинами. Волотные почвы-преимущественно атмосферного увлажнения (торфяники верхового и переходного типов). Подзолистые почвы
таежной зоны европейской части СССР неодинаковы: для северной подзоны характерны подзолистые гумусо-железистые
и подзолисто-глееватые почвы, для южной подзоны-дерново-
подзолистые. В таежной зоне по мере продвижения с севера
Ha юг в лесных почвах уменьшается содержание грубых скелетных частиц, ослабляется процесс оглеения и усиливается
дерновый. Почвы становятся тяжелее по механическому составу, увеличивается содержание подвижных форм, усиливаются процессы гумификации и нитрификации и т. п. Иными
словами, к югу увеличивается производительность почв.
Определение типа леса включает B себя как лесораститель-
ные условия того или иного участка лесной территории, так
и сам древесный фитоценоз. Многочисленные исследования показали, что в определенных узких географических условиях
тип леса характеризуется своей, " присущей ему почвой.
Почва-один из ведущих компонентов биогеоценоза. На изучение взаимосвязей почвы и фитоценоза необхОдимо обращать
самое серьезное внимание (Зонн, 1967). Лесные фитоценозы
играют и определенную "почвосозидающую роль, различающуюся для типов лесных биогеоценозов. Это проявляется
в различных для каждого типа леса качествах и количествах
древесного и нижнего ярусов опада, условиях минерализации,
скорости ее и в соотношении процессов элювиальных и акку-
мулятивных.
Основные хвойные породы тайги (Сосна, ель и лиственница)
произрастают на почвах самого разнообразного механического
состава, от глубоких песков до глин. Сосна произрастает на
почвах разного механического состава, на тяжелых почвах она
образует более продуктивные древостои. Однако как светолюбивая порода она не выдерживает в этих условиях конкуренции с елью и при отсутствии пожаров или других катастрофических сукцессий уступает место этой теневыносливой породе.
Успешно сохраняет свои позиции сосна на сухих легких почвах, особенно в южной половине таежной зоны. Ель также
произрастает на почвах разного механического состава.
На легких свежих почвах она успешно растет в северной
и средней тайге. На тяжелых (суглинистых и глинистых) почвах ель растет повсеместно. Лиственница в пределах современного ареала произрастает на разных почвах. Чаще она
встречается на почвах, сформированных на близко залегающей
известковой плите или гипсах, а также почвах, сформированных на известняковистой морене. Пихта занимает те же почвы,
что и ель.
Еловые древостои также формируются и произрастают
в различных лесорастительных условиях, отмечающихся по
подзонам тайги. На рис. 17 показаны ареалы распространения
еловых лесов (типов леса), нанесенные на ординатную сетку
Сукачева. В предтундровых лесах ель произрастает B различных лесорастительных условиях от лишайниковых до логовоприручейных типов и от “сфагновых до черничннков. По мере
продвижения на юг ареал ельников приближается к брусничникам и сложным. В таежной зоне европейской части России
ПОЧВЫ еловых лесов характеризуются ВЛАЖНОСТЬЮ, достаточной ДЛЯ успешного роста древесных пород, здесь нет излишне
сухих почв. Но в этих условиях в некоторых типах леса наблюдается сезонный или постоянный избыток влаги.
 Рис.
17. Ареалы некоторых типов ельников в таежной зоне:
Рис.
17. Ареалы некоторых типов ельников в таежной зоне:
1-лишайннковыи; {2-брусничник; з-черничник свежий; 4-рябинниковый; 5-кис-
личник: б-сложный; 7-крупнотравный; 8-долгомошник; 9-кустарннчково-сфагно-
вый; 10-сфагновый; 11-травяно-сфагновый; 12-логово-приручейный
показателем водно-воздушного режима ПОЧВ может служить отношение мощности органического горизонта ho к общей
МОЩНОСТИ ПОЧВЫ Н, выраженное в ДОЛЯХ единицы.
Наши исследования показали, что при благоприятном вед-
НОМ режиме и хорошей аэрации отношение органического ГО°
ризонта к общей глубине почвы составляет 0,1-0,2. По мере ухудшения аэрации и увеличения влажности почвы отношение
возрастает до 1.
Водно-воздушный режим ho : Н:
благоприятный . . . . . . . . . . . . . О,1-0,2;
затрудненный . . . . . . . . . . . . .0,25-0,4;
плохой.................0,5-0,7;
очень плохой . . . . . . . . . . . . . . 0,8-1‚0.
Это связано с ухудшением условий минерализации лесного
опада и накоплением органики в условиях затрудненного дренажа и избытка влаги.
На почвах с избыточным увлажнением формируются леса
пониженной продуктивности. Избыток влаги в условиях тайги
приведит к более позднему оттаиванию почвы, неблагоприятному водно-воздушному режиму, малому содержанию или отсутствию в переувлажненных горизонтах почвы кислорода. 50-
лотно-подзолистые и болотные почвы характеризуются также
высокой кислотностью, низкой степенью насыщенности основаниями. В этих условиях у древесных пород развивается поверх-
ностная корневая система. Корнеобитаемый слой почвы имеет
незначительную мощность (20-30 см) и невысокие запасы
питательных веществ. Продуктивность древостоев в этих усло-
виях низкая, класс бонитета обычно V. Наибольшая продуктив-
ность в таежной зоне отмечается у древостоев на почвах сред-
него увлажнения (свежие почвы).
Лесная растительность в свою очередь сильно влияет на
почву H почвенные процессы. Особенно это влияние сказы-
вается через опад. Масса ежегодно отмирающих частей расте-
ний велика и составляет в северной подзоне тайги в ельнике-
черничнике свежем в абсолютно сухом состоянии 1,7-2,5 т
(Паршевников, Черных, 1968). В средней подзоне тайги сред-
неголрвое количество опада в спелых ельниках-зеленомошни-
ках составляет 2,2-2,6 т/га (Паршевников, 1962; Егорова,
1968). В южной тайге (Смирнов, 1971) масса опада составляет
2,9 T B ельнике-кисличнике (75 лет), 3,5 т в ельнике кислично-
папоротниковом. Масса опада в южнотаежных ельниках кис-
лично-папоротниковом и кислично-черничном равна 2,4 T,
B черничнике 1,8 т/га (Кошельников, 1964). Содержание золь-
ных элементов и химический состав золы у ели, а также опада
и подстилок зависит от химического состава почвы (Морозова,
Куликова, 1968; Паршевников, Черных, 1968). Следовательно,
будет разница и по лесорастительным регионам тайги. Ежегодно в почву с опадом древесных порол поступает значитель-
ное количество минеральных веществ.
Почвоулучшающая роль наиболее выражена у лиственных
пор0д (Ткаченко, 1952; Степанов, 1932). Березняки, формирующиеся на месте ельников, вызывают большую интенсификацию
биологического круговорота веществ, ослабляют подзолообразовательный процесс (Фролова, 1968). Опад ели в меньшей
степени, чем опад сосны, обогащает почву азотом, калием
и марганцем и B большей-кальцием (Тонконогов, 1968).
Не менее важный источник поступления органического вещества B почву-отмершие корни деревьев. Ежегодно отми-
рающие корни представляют заметный вес в _общем поступлении органики в почву.
Гумификация растительных остатков B подстилке и мине-
ральных горизонтах почвы приводит к обогащению почвы ор-
ганикой. В условиях выраженного промывного режима под-
золистых почв значительные количества органики вмываются
B минеральные горизонты из псдстилки. Количество органиче-
ского вещества‘ B подзолистых почвах ельников-черничников
достигает 8,3-13,1 кг на 1 м"2 профиля (табл. 23). На 1 м2
почвенного профиля содержится более 200 г воднораствори-
мого подвижного органического вещества.
Проводятся исследования с целью определения непосредственного влия-
ния живых растении на почву. Многие ученые признают лишь косвенное
влияние путем воздействия ПРОДУКТОВ распада растительного вещества на
почву. В. И. Вернадский (1954) допускал возможность непосредственного
влияния жизни на химические элементы земной коры. А. А. Роле (1944)
высказал предположение, что в подзолообразовании принимают участие
ионы водорода, продуцируемые живыми корнями растений. Впоследствии
это предположение поддержали И. Н. Гоголев (1968) и др. Эта теория
еще не получила дОСТЗТОЧНОГО подтверждения И является СПОРНОЙ.
Лес значительно влияет на климат почвы, а черезнего на
течение ряда почвенных и биологических процессов. Характерная черта почвообразования в таежной зоне-интенсивное
накопление органического вещества на поверхности почвы
в виде лесной подстилки или торфянистого (торфяного) горизонта. Этот органический горизонт и является основным хра-
нителем и источником питательных веществ для древесной растительности, подавляющая часть корневой системы которой
расположена в ‚подстилке. По мнению В. В. Пономаревой
(1964), «это приспособительная реакция организмов к сильно
выраженным элювиальным условиям, при которых невозможно
сохранение накопленных элементов питания вне живых органов растений и их отмирающих остатков» (с; 331). При этом следует учитывать, что еловый лес всегда стремится развить корневую систему вглубь. На легких супесчаных почвах корни ели проникают в минеральные горизонты на глубину 3 M. Правда,
это меньшая часть корней (до 20% массы), но она-токак раз
и служит основой получения дополнительного количества минеральных элементов. С годовым опадом они попадают в подстилку и вступают в активный уже биологический круговорот
веществ. При кажущейся незначительности этого биологического транспорта минеральных веществ за период; измеряемый
тысячами лет‚ он достигает значительных величин. Иными словами, лесная растительность, в частности еловая, таким путем
создает специальный органический горизонт"почвы, где образуются значительные запасы питательных веществ и где лесные породы, в основном микотрофные, находят оптимальные условия для развития корневых систем, интенсивно питающих
весь организм.
В таежной зоне бывают годы, когда в минеральных горизонтах весь вегетационный период или часть его создаются
неблагоприятные условия для деятельности корневых систем
(избыток влаги, или низкие температуры). Подстилка же является тем страховым горизонтом, где всегда может функционировать корневая система.
При биологической переработке органического вещества
В почве также выделяется и тепло, т. е. подстилка В какой-то
мере сама отепляется. При этом выделяется углекислый газ,
который идет на фотосинтетические процессы в данном биогеоценозе. Количество углекислоты даже в северной тайге
в наиболее теплый период превышает 300 мг/ч с 1 м2 (Пар-
шевннков, 1969). Роль лесной подстилки, как наиболее благоприятного почвенного горизонта, возрастает С юга на север.
По мере продвижения на север, помимо выраженной элю-
внальности почв, большую роль начинает играть и фактор
тепла. Подстилка является горизонтом ПОЧВЫ, характеризующимся наиболее длительным теплым периолом и наиболее
ВЫСОКИМИ температурами В слое ПОЧВЫ, ДОСТУПНОМ КОРНЯМ де-
ревьев. Таким образом, лес сам создает для свой жизнедея-
тельности почвенный горизонт (подстилку), обладающий наи-
более благоприятными лесорастительными условиями ‚и В ТОМ
числе питательным режимом.
В МОЩНОСТИ подстилки, в аккумуляции элементов пищи
в ней, наблюдается определенная географическая закономер-
ность. По мере продвижения с юга таежной зоны на север,
т. е. по мере продвижения древесной растительности В менее
благоприятные условия, увеличиваются мощность и запас орга-
нического вещества в виде лесной подстилки.
Верхний, слаборазложившийся слой подстилки характеризуется значительным соцержанием кальция, калия, магния.
С увеличением разложенности подстилки содержание указанных элементов снижается, увеличивается содержание кремне-
кислоты, железа и алюминия (Егорова, 1968). Содержание
минеральных элементов в течение года непостоянно. Наиболее
выражена динамичность подвижных форм элементов питания
растений в подстилке. Максимальное содержание подвижных
элементов отмечается весной и осенью, минимальное-в се-
редине вегетационного перИОДа (Куликова, 1968). Это может
быть в какой-то степени объяснено сильным потреблением рас-тениями минеральных элементов и азота в период новообразования органов и создания запасов на следующий год. Более углубленное изучение лесных подстилок показало значительные различия и в химическом составе их в различных типах
леса (Чернов, 1965; Левина, 1960; Левкина и Яковлев, 1965;
Куликова, Егорова, 1965; Перевозчикова, Пахнутова, 1965).
Запас и свойства лесных подстилок тесно связаны с типом
леса, т. е. количеством и составом лесного опада. Лесные подстилки влияют на формирование профиля лесных почв. Вымываемые при разложении подстилки воднорастворимые вещества способствуют увеличению подвижности железа (Зонн,
1964; Кауричев, 1964; Кошельков, 1964; Сапожников, 1968;
Фролова, 1968). Наибольшая восстановительная способность
подстилок отмечается в начале и конце вегетационного периода (Сапожников, 1968; Фролова, 1968). Количество водно-растворимого органического вещества (в пересчете на углерод), поступающего из подстилки ельника-черничннка за ве-
гетационный период (май-сентябрь), составляет 150-
200 кг/га, из подзолистого горизонта - 80 кг/га.
Таким образом, лес, создавая специфическую, свойственную
тому или иному типу леса подстилку, влияет и на свойства
нижележащих горизонтов почвы.
Кислотность почвы
Кислотность почвы формируется псд влиянием двух основ-
ных факторов-химического состава древесного опада и химического состава порол,
на которых формируется почва. В зависимости от минералогического состава подстилающие породы могут быть кислыми, нейтральными или щелочными.
Реакция подстилок и верхних горизонтов почвы в значительной степени обусловливается составом и характером разложения опада. Как правило, под таежными лесами верхние
горизонты почвы, сформировавшиеся как на кислых, так и щелочных подстилающих породах, имеют кислую реакцию. Это,
вероятно, является своеобразной защитной реакцией леса против наступления травянистой, в том числе степной, растительности. В процессе эволюции лесная растительность выработала
способность успешного роста при кислой или слабокислой
реакции почвы. Так, по исследованиям А. Ф. Иванова (1970),
успешный рост пихты сибирской наблюдается при рН в0дной
вытяжки 4,5-5‚6; ели обыкновенной-5,2-6,6; сосны обык-
новенной - 5-62; лиственницы сибирской - 5,7-6,2.
Таежная древесная растительность сама создает наиболее
оптимальную кислую среду верхних горизонтов почвы. В первую очередь кислотность подстилки может быть объяснена
реакцией сока хвои. По исследованиям, проведенным
А. Ф. Ивановым, рН сока хвои ели обыкновенной равен 3-3,5;
лиственницы сибирской-3,3-4. Влияние ели, пихты и лист-
венницы на кислотность почвы прослеживается во всех горизонтах почвы (Иванов, 1970). Сосна увеличивает кислотность
преимущественно в верхнем горизонте почвы.
В результате вырубки хвойного леса временно разрастается травянистая растительность, вызывающая изменение реакции
почвы.
Напочвенный покров на вырубках 4-летней давности влияет на изме-
нение КИСЛОТНОСТИ ТОЛЬКО подстилки. ЭТИ изменения не ПРОЯВЛЯЮТСЯ В ПОДЗОЛИСТОЙ
и более глубоко лежащих горизонтах почвы (табл. 26).
Наименьшая кислотность (6,1) верхней части подстилки наблюдается под иван-чаем, где реакция близка к нейтральной,
затем под вейником (5,5). Отпад кустарничков и зеленых мхов дает наиболее кислую реакцию. Дожцевые осадки, стекающие по стволам ели, также имеют кислую реакцию. Кислотность осадков пед кронами ели может достигать 5 (Куликова, 1968
и др.).
Анализ кислотности верхних горизонтов почвы в различных типах еловых лесов показал четкую взаимосвязь (табл. 27).


OT шейки корня отходит вниз один корень, который в дальнейшем начинает давать боковые ответвления. На второй год жизни вертикальный тип корневой системы сохраняется, но на
строении корневой системы уже начинают сказываться лесорастительные условия. На супесчаных, хорошо дренированных почвах центральный вертикальный корень продолжает рост, но интенсивность его уменьшается. Начиная со 2 года жизни,
у ели интенсивно растут боковые корни (5-8 шт.). Общая
протяженность их (исключая сосущие корни) к возрасту 5 лет
достигает 100 см, в 10-12 лет-360 см. Корневые системы ели
в условиях хорошо дренированных супесчаных почв характе-
ризуются выраженной мочковатостью, на 1 см проводящих
корней приходится 5 сосущих корней 1 порядка (табл. 28).
Все это свидетельствует о большой роли растительности
тайги в создании наиболее благоприятных условий кислот-
ности почвы для своего роста и одновременно о большом зна-
чении подкисляющей роли древесных пород для удержания
позиций тайги в ее взаимоотношении с травянистыми форма-
циями.
Почва и корневая система
Корневая система растений помимо чисто механических
функций прикрепления растения к почве выполняет рЯД дру-
гих, более важных. При помощи корней растение поглощает
из почвы влагу и растворимые минеральные вещества, прово-
дит и частично подает их в стебель или ствол. Корни участ-
вуют и в обеспечении растения углекислотой (Курсанов, 1954,
1957). Корневая система является также и активной частью
растения, в ней происходит биосинтез разнообразных органи-
ческих соединений (Сабинин, 1949; Шмук, 1941; Курсанов,
1954; Кретович и др., 1959). Корневая система растений, по-
добно грибам и бактериям, обладает способностью разлагать
сложные органические соединения при" помощи выделяемых
ферментов. При определенных условиях питания, а может быть и всегда, корни могут и усваивать эти продукты (Красильни-
ков,1952).
Роль корневой системы в жизни растения очень важна и разнообразна.
Но корневые системы лесных древесных пород изучены недостаточно. В той
или иной степени они характеризуются в работах Н. А. Качинского (1925),
И. С. Мелехова (1957, 1960), А. П. Шиманюка (1950), А. П. Тыртикова
(1954), A. я. Орлова (1957, 1960) и др.
Почвенные условия сильно влияют на характер и строение
корневых систем древесных растений.
В условиях европейской тайги можно выделить три кате-
гории корневых систем:
1. Глубокая корневая система-формируется на Глубоких
песчаных или супесчаных, достаточно увлажненных почвах.
Корни такой ели проникают до 3-4 м.
2. Якорная корневая система-формируется на двучлен-
ных дренированных почвах. Верхний член супесчаный, мощность его 0,5-1 м. Корневая система представлена сочетанием поверхностной корневой системы с якорными корнями.
3. Поверхностная корневая система-формируется на тяжелых по механическому составу почвах, а также на избыточно увлажненных и болотных почвах. Корневая система
представлена хорошо развитыми корневыми лапами и корнями,
идущими горизонтально и преимущественно в подстилке.
У ели в первый год жизни корневая система строится по
вертикальному типу не зависимо от условий произрастания.
Oт шейки корня отходит вниз один корень, который B дальнейшем начинает давать боковые ответвления. На второй год
жизни вертикальный тип корневой системы сохраняется, но на
строении корневой системы уже начинают сказываться лесо-
растительные условия. На супесчаных, хорошо дренированных
почвах центральный вертикальный корень продолжает рост,
но интенсивность его уменьшается. Начиная со 2 года жизни,
у ели интенсивно растут боковые корни (5-8 шт.). Общая протяженность их (исключая сосущие корни) к возрасту 5 лет
достигает 100 см, в 10-12 лет-360 см. Корневые системы ели
в условиях хорошо дренированных супесчаных почв характеризуются выраженной мочковатостью, на 1 см проводящих корней приходится 5 сосущих корней 1 порядка (табл. 28).

В дальнейшем, в зависимости от почвенных условий форми-
руется или якорная, или глубокая корневая система. В ельниках-черничниках на слабопедзолистой гумусово-железистой
песчаной почве., y ели наряду с поверхностными далеко иду-
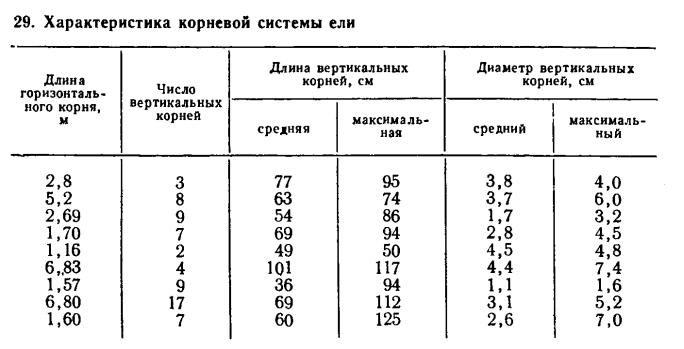
щими корнями имеются вертикальные якорные корни, проникающие на глубину до 1 м (табл. 29).
Наибольшее количество корней (на площади 0,1 м2 в каждом горизонте) наблюдается на расстоянии 1 м от дерева, где
насчитывается до 593 крупных (диаметром более 3 M) и мелких корней (табл. 30). Наибольшее число корней, преимущественно мелких, наблюдается B гумусово-железистом горизонте
В,. В подстилке, где сосредоточено около 75% общего количества корней, преобладают (25%) крупные корни. Масса корней уменьшается с глубиной, исключение составляет горизонт
B1 (табл. 31), B котором отмечается наибольший удельный вес
деятельных корней (около 25%). Беден мелкими корнями подзолистый горизонт. В ельниках-зеленомошниках чистых и в
брусничниках при произрастании на глубоких песках ель фор-
мирует глубокую корневую систему. Стержневой корень ее проникает вглубь до 3 M, до влажных подстилающих горизонтов.
Отмечается также сильное ветвление корней в горизонте В1.
B условиях временного или постоянного избытка влаги у ели наблюдаются особенности в формировании корневых систем. Эти особенности в основном заключаются в придаточном корнеобразовании. Анаэробные условия в минеральных горизонтах и нижней части подстилки в течение весны, начала лета
и осенью вызывают отмирание растущих здесь корней ели
и других пород. Одновременно на стволе деревца (выше корневой шейки), по мере его зарастания влажным мхом, появляются придаточные корни. Многие древесные породы только
потому и выживают в период своего возобновления, что способны образовывать на стебле придаточные корни. Причины, вызывающие образование придаточных корней, окончательно
еще не выявлены. Для `образования придаточных корней нужна изоляция от света. B сухой почве корни не образуются. Общим условием является достаточная аэрация и необходимая температура почвы (Кощеев, 1955).

Ha ТОРФЯНИСТО-ПОДЗОЛИСТЫХ почвах (ельники-черничники влажные, долгомошники и т. п.) y ели на 2-й год жизни вер-
тикальный тип корневой системы сохраняется, но рост верти-
кального корня задерживается. В наиболее увлажненных местах отмечено даже загнивание нижней части его. Рост боковых корней хороший. Они распространяются в стороны до 4см.
В З-летнем возрасте наиболее интенсивно растут боковые
корни. Рост вертикального корня прекращается. Вследствие
плохих условий аэрации B нижней части торфянистого горизонта, а также благодаря окружению стебля влажным моховым очесом начинают образовываться придаточные корни.
В этом возрасте придаточные корни имеются у 5-6% елочек.
У 4-летних елочек придаточные корни обнаружены у 17% деревцев. У них корневая ‘система больше распространена в стороны, чем вглубь. Корни проникают на глубину 6-7 см, в стороны они идут на 9-10 см. В возрасте 7-10 лет происходит дальнейшая замена основных (первичных) корней придаточными. В этом возрасте первичные корни растут очень мед-
ленно, часть из них отмирает. Это вызывается недостатком кислорода в нижних частях торфянистой подстилки, увеличением ее мощности над корневой системой. У ели этого возраста придаточные корни отсутствуют примерно только у 10%
деревец. Здесь еще сильнее проявляется горизонтальное распространение корней. Корни 7-10-летнего самосева ели расходятся в стороны на 20-35 см, B отдельных случаях до 45 см и более. Глубина же проникновения корней обычно не превышает 8-10 см. Изредка встречаются еще более плоские корневые системы (глубиной 3-5 CM).
У подроста ели B возрасте 15-25 лет и более наблюдается
полная замена первичной корневой системы. Место первичных корней занимают придаточные корни, появляющиеся выше по
стволу в верхней части торфянистого горизонта, где кислородный режим более благоприятен для роста. Наивысший для ели возраст, в котором еще могут появляться адвентивные корни,
исследователями не указывается. Нами встречена ель в возрасте 115-120 лет, возраст придаточного корня у которой
20-25 лет. Следовательно, он появился в 90-100-летнем воз-.
расте дерева.
Формирование корневой системы за счет придаточных кор-
ней прив0дит к концентрации корней в верхних горизонтах.
Только B подстилке располагается около 75% массы корневой
системы . В результате различий в строении корне-
вых систем ели, вызванных почвенными условиями, меняется и насыщенность почвы корнями.
В типах леса на заболоченных почвах насыщенность почвы
корнями несколько меньше, чем B типах на суходольных поч-
вах.

Главное различие между ними заключается в распределении корней no почвенному профилю. Если на суходольных
почвах корни в заметных количествах (10-15% no массе) встречаются в нижнем почвенном горизонте B, то на торфянисто-подзолистых почвах корни в горизонте В отсутствуют.
Заключение
Еловые леса приурочены к зоне прохладного и влажного климата, гидротермический коэффициент изменяется от 0,57 до 0,38. Показатель увлажнения высокий и составляет по подзонам тайги 0,7-0,6. Радиационный индекс сухости колеблется от 0,85 до 1. Это зона четко выраженных времен года с холодной снежной зимой и умеренно теплым, с достаточным количеством осадков летом. Количество осадков превышает потенциальную возможность их испарения. Влажность воздуха достаточно высокая.
Еловые леса произрастают преимущественно на подзолистых почвах, умеренно теплых и достаточно увлажненных в течение всего вегетационного периода.
Еловые леса заметно влияют на климат и почву, трансформируют их в благоприятном для себя направлении как с точки зрения обеспечения энергии роста. Так и с точки зрения конкурентных отношений с другими видами растений, за выживаемость ели как вида.
Библиографический список
1. ЛевкинаТ. И., Яковлев Ф. С. Взаимоотношения растительности и почв
в ельниках заповедника ‹Кивач».-В кн.: Плодородие почв Карелин. М.-Л.,
«Наука», 1965, с. 5-19.
2. Пьявченко Н. И. Лесное болотоведение. М., изд-во АН СССР, 1963.
191 с.
3. Пьявченко Н. И., Сибирева 3. А. О роли атмосферной пыли в питании
болот.- «Доклады АН СССР», 1959, т. 124, № 2, с. 414-417.
4. Пьявченко Н. И. Типологическая характеристика заболоченных лесов
европейской части РСФСР применительно к задачам осушения-В кн.: Про-
блемы повышения продуктивности лесов. М.-Л.‚ Гослесбумиздат, 1959, т. 2,
с. 7-4].
5. Пятницкий Г., К. Материалы к познанию короедов ельников Пошехон-
ского уезда Ярославской губернии.- «Защита растений», 1930, т. VI, № 5-6,
с. 595-630.
6. Разумовская 3. Г., Мустафова Н. Н. О биологической активности почв
ельника-кисличника и ельника-черничника.- ‹Вестник ЛГУ», 1959, вып. 1,
N9 3, с. 48-55.
7. Райд Л.В. O взаимосвязях между влажностью почвы, метеорологическими
условиями и грунтовыми водами в некоторых типах леса.-_ «Биология», 1968,
17, № 3, с. 313-319.
8. Paнe А. А. O возможной роли растительности в подзолообразовании
(предварительное сообщение).-_ «Почвоведение», 1944, № 4-5, с. 159-179.
Сабинин Д. A. O значении корневой системы в жизнедеятельности расте-
ний.- «Тимирязевские чтения», IX. М., изд-во АН СССР, 1949. 48 с.
9. Сапожникова С. A. Опыт агроклиматического районирования территории
СССР.-В кн.: Вопросы агроклиматического районирования СССР. М., изд.
МСХ СССР, 1958, с. 14-37.
10. Сахаров М. И. О влиянии отдельных ярусов лесных ценозов на радиацию и освещенность.- «Доклады AH СССР», 1948, т. 62, № 5, с. 701-704.
11. Сахаров M. И. Промерзание и размерзаине почв в лесных фитоценозах.-
‹Метеорология и гидрология», 1938, N9 11-12, с. 136-148.
 (zip - application/zip)
(zip - application/zip)