1.Протопласт растительной клетки, его микроскопические и ультрамикроскопические структурные элементы (цитоплазма, ядро, пластиды, митохондрии, рибосомы и др.)
Клетка - основная форма организации живой материи, элементарная единица организма. Она представляет собой самовоспроизводящуюся систему, которая обособлена от внешней среды и сохраняет определенную концентрацию химических веществ, но одновременно осуществляет постоянный обмен со средой. Клетка - основная структурная единица одноклеточных, колониальных и многоклеточных организмов. Единственная клетка одноклеточного организма универсальна, она выполняет все функции, необходимые для обеспечения жизни и размножения. У многоклеточных организмов клетки чрезвычайно разнообразны по размеру, форме и внутреннему строению. Это разнообразие связано с разделением функций.
Протопласт – активное живое содержимое клетки. Протопласт представляет собой чрезвычайно сложное образование.
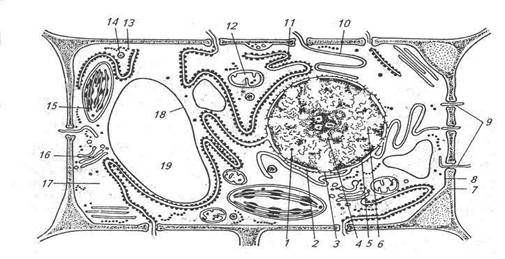
Строение растительной клетки по данным электронной микроскопии: 1 – ядро; 2 – ядерная оболочка; 3 – ядерная пора; 4 – ядрышко; 5 – хроматин; 6 – кариоплазма; 7 – клеточная стенка; 8 – плазмалемма; 9 – плазмодесмы; 10 – агранулярная эндоплазматическая сеть; 11 – гранулярная эндоплазматическая сеть; 12 – митохондрия; 13 – рибосомы; 14 – лизосома; 15 – хлоропласт; 16 – диктиосома; 17 – гиалоплазма; 18 – тонопласт; 19 – вакуоль.
Химический состав протопласта очень сложен и разнообразен. Каждая клетка характеризуется своим химическим составом в зависимости от физиологических функций. Основными классами соединений являются: вода (60-90%), белки (40-50% сухой массы протопласта), нуклеиновые кислоты (1-2%), липиды (2-3%), углеводы и другие органические соединения. В состав протопласта входят и неорганические вещества в виде ионов минеральных солей (2-6%). Белки, нуклеиновые кислоты, липиды и углеводы синтезируются самим протопластом.
Цитоплазма -обязательная часть живой клетки, где происходят все процессы клеточного обмена, кроме синтеза нуклеиновых кислот, совершающегося в ядре. Основу цитоплазмы составляет ее матрикс, или гиалоплазма,в который погружены органеллы. Органелла — это постоянное образование в клетке, которое выполняет определенные функции. Их еще называют органоидами. Органелла — это то, что позволяет клетке жить. Точно так же, как животное и человек состоят из органов, так каждая клетка состоит из органоидов. Они разнообразны и выполняют все функции, обеспечивающие жизнь клетки: это и обмен веществ, и их запасание, и деление.
Гиалоплазма – сложная бесцветная, оптически прозрачная коллоидная система, она связывает все погруженные в нее органеллы, обеспечивая их взаимодействие. Гиалоплазма содержит ферменты и активно участвует в клеточном метаболизме, в ней протекают такие биохимические процессы, как гликолиз, синтез аминокислот, синтез жирных кислот и масел и др. Она способна к активному движению и участвует во внутриклеточном транспорте веществ.
Мембраны – живые компоненты цитоплазмы. Они отграничивают протопласт от внеклеточной среды, создают внешнюю границу органелл и участвуют в создании их внутренней структуры, во многом являясь носителем их функций. Характерной особенностью мембран является их замкнутость, непрерывность – концы их никогда не бывают открытыми. В некоторых особенно активных клетках мембраны могут составлять до 90% сухого вещества цитоплазмы.
Пограничными мембранами протопласта являются плазмалемма – плазматическая мембрана и тонопласт – вакуолярная мембрана. Плазмалемма – наружная, поверхностная мембрана цитоплазмы, обычно плотно прилегает к клеточной стенке. Она регулирует обмен веществ клетки с окружающей средой, воспринимает раздражения и гормональные стимулы, координирует синтез и сборку целлюлозных микрофибрилл клеточной стенки. Тонопласт регулирует обмен веществ между протопластом и клеточным соком.
Митохондрии - органоиды обеспечивают клетку энергией — они отвечают за клеточное дыхание. Они есть и у растений, и у животных, и у грибов. Данные клеточные органеллы имеют две мембраны: внешнюю и внутреннюю, между которыми есть межмембранное пространство. То, что находится внутри оболочек, называется матриксом. В нем находятся разнообразные ферменты — вещества, необходимые для ускорения химических реакций. Внутренняя мембрана обладает складками — кристами. Именно на них и происходит процесс клеточного дыхания. Кроме того, в матриксе митохондрий находится митохондриальная ДНК (мДНК) и мРНК, а также рибосомы, практически аналогичные тем, которыми обладают прокариотические клетки.
Рибосома- органоид отвечает за процесс трансляции, при котором из отдельных аминокислот синтезируется белок. Строение органеллы рибосомы проще, чем митохондрии, — она не обладает мембранами. Данный органоид состоит из двух частей (субъединиц) — малой и большой. Когда рибосома бездействует, они находятся раздельно, а когда она начинает синтезировать белок — объединяются. Также собираться вместе могут и несколько рибосом, если полипептидная цепочка, синтезируемая ими, очень длинная. Такая структура называется "полирибосома".
Рибосома состоит из двух субъединиц (большой и малой), соединенных между собой ионами магния. Субъединицы образуются в ядре, а именно в ядрышке, сборка рибосом осуществляется в цитоплазме.
Эндоплазматическая сеть (эндоплазматический ретикулум) представляет собой разветвленную трехмерную сеть каналов, пузырьков и цистерн, ограниченных мембранами, пронизывающую гиалоплазму. Эндоплазматическая сеть в клетках, синтезирующих белки, состоит из мембран, несущих на наружной поверхности рибосомы. Такая форма получила название гранулярной, или шероховатой . Эндоплазматическая сеть, не имеющая рибосом, называется агранулярной, или гладкой. Агранулярная эндоплазматическая сеть принимает участие в синтезе жиров и других липофильных соединений (эфирные масла, смолы, каучук).
Эндоплазматическая сеть функционирует как коммуникационная система клетки и используется для транспортировки веществ. Эндоплазматические сети соседних клеток соединяются через цитоплазматические тяжи – плазмодесмы, которые проходят сквозь клеточные стенки. Эндоплазматическая сеть – центр образования и роста клеточных мембран. Она дает начало таким компонентам клетки, как вакуоли, лизосомы, диктиосомы, микротельца. При посредстве эндоплазматической сети осуществляется взаимодействие между органеллами.
Аппарат Гольджи названпоимениитальянскогоученого К. Гольджи, Это комплекс дискообразных мембранных мешочков, называемых цистернами. Функции этого органоида заключаются в сортировке, запасании и превращении некоторых веществ. Синтезируются здесь в основном углеводы, которые входят в состав гликокаликса
Лизосомы Функции органелл этого вида сводятся к осуществлению клеточного пищеварения. Лизосомы обладают одной мембраной, внутри которой находятся ферменты — катализаторы химических реакций. Иногда эти органоиды не только расщепляют питательные вещества, но и переваривают целые органоиды. Такое может происходить при длительной голодовке клетки и позволяет ей жить еще некоторое время. Хотя если питательные вещества все еще не начнут поступать, клетка умирает.
Пластиды –органеллы, характерные только для растений. Различают три типа пластид: 1) хлоропласты (пластиды зеленого цвета); 2) хромопласты (пластиды желтого, оранжевого или красного цвета) и лейкопласты (бесцветные пластиды). Обычно в клетке встречаются пластиды только одного типа.
Хлоропласты имеют наибольшее значение, в них протекает фотосинтез, образование органических веществ из неорганических за счет энергии света Они содержат зеленый пигмент хлорофилл, придающий растениям зеленый цвет, и пигменты, относящиеся к группе каротиноидов. Каротиноиды имеют окраску от желтой и оранжевой до красной и коричневой, но обычно она маскируется хлорофиллом. Каротиноиды делят на каротины, имеющие оранжевую окраску, и ксантофиллы, имеющие желтую окраску.
Лейкопласты - мелкие бесцветные пластиды. Они встречаются в основном в клетках органов, скрытых от солнечного света, таких как корни, корневища, клубни, семена.
Хромопласты - пластиды оранжевого, красного и желтого цвета, который обусловлен пигментами, относящимися к группе каротиноидов. Хромопласты встречаются в клетках лепестков многих растений (ноготки, лютик, одуванчик), зрелых плодов (томат, шиповник, рябина, тыква, арбуз), редко - корнеплодов (морковь), а также в осенних листьях.
Ядро – основная и обязательная часть эукариотической клетки. Ядро является центром управления обменом веществ клетки, ее ростом и развитием, контролирует деятельность всех других органелл. Ядро хранит генетическую информацию и передает ее дочерним клеткам в процессе клеточного деления. Ядро имеется во всех живых растительных клетках, исключение составляют только зрелые членики ситовидных трубок флоэмы. Клетки с удаленным ядром, как правило, быстро погибают.
Ядро – самая крупная органелла, его размер составляет 10-25 мкм. Очень большие ядра у половых клеток (до 500 мкм). Форма ядра чаще сферическая или эллипсоидальная, но в сильно удлиненных клетках может быть линзовидной или веретеновидной.
Клетка, как правило, содержит одно ядро. В молодых (меристематических) клетках оно обычно занимает центральное положение. По мере роста центральной вакуоли ядро смещается к клеточной стенке и располагается в постенном слое цитоплазмы.
По химическому составу ядро резко отличается от остальных органелл высоким (15-30%) содержанием ДНК – вещества наследственности клетки. В ядре сосредоточено 99% ДНК клетки, она образует с ядерными белками комплексы – дезоксирибонуклеопротеиды. В ядре содержатся также в значительных количествах РНК (в основном иРНК и рРНК) и белки.
Структура ядра одинакова у всех эукариотических клеток. В ядре различают хроматин и ядрышко, которые погружены в кариоплазму; от цитоплазмы ядро отделено ядерной оболочкой с порами.
Ядерная оболочка состоит из двух мембран. Наружная мембрана, граничащая с гиалоплазмой, несет прикрепленные рибосомы. Оболочка пронизана довольно крупными порами, благодаря которым обмен между цитоплазмой и ядром значительно облегчен; через поры проходят макромолекулы белка, рибонуклеопротеиды, субъединицы рибосом и др. Наружная ядерная мембрана в некоторых местах объединяется с эндоплазматической сетью.
Кариоплазма (нуклеоплазма, или ядерный сок) – основное вещество ядра, служит средой для распределения структурных компонентов – хроматина и ядрышка. В ней содержатся ферменты, свободные нуклеотиды, аминокислоты, иРНК, тРНК, продукты жизнедеятельности хромосом и ядрышка.
Ядрышко - плотное, сферическое тельце диаметром 1-3 мкм. Обычно в ядре содержатся 1-2, иногда несколько ядрышек. Ядрышки являются основным носителем РНК ядра, состоят из рибонуклеопротеидов. Функция ядрышек – синтез рРНК и образование субъединиц рибосом.
Хроматин - важнейшая часть ядра. Хроматин состоит из молекул ДНК, связанных с белками, - дезоксирибонуклеопротеидов. Во время деления клетки хроматин дифференцируется в хромосомы. Хромосомы представляют собой уплотненные спирализованные нити хроматина, они хорошо различимы в метафазе митоза, когда можно подсчитать число хромосом и рассмотреть их форму. Хроматин и хромосомы обеспечивают хранение наследственной информации, ее удвоение и передачу из клетки в клетку.
Число хромосом у разных организмов варьирует от двух до нескольких сотен. Как правило, каждый вид имеет характерный и постоянный набор хромосом, закрепленный в процессе эволюции данного вида. Изменение хромосомного набора происходит только в результате хромосомных и геномных мутаций. Наследственное кратное увеличение числа наборов хромосом получило название полиплоидии, некратное изменение хромосомного набора – анеуплоидии. Растения – полиплоиды характеризуются более крупными размерами, большей продуктивностью, устойчивостью к неблагоприятным факторам внешней среды. Они представляют большой интерес как исходный материал для селекции и создания высокопродуктивных сортов культурных растений. Полиплоидия также играет большую роль в видообразовании у растений.
2.Оболочка растительной клетки, ее химический состав, строение и вторичные изменения (одревесение, опробкование и др.)
Прочная оболочка — характерный признак растительной клетки, отличающий ее от клетки животной. Оболочка придает клетке определенную форму и прочность и защищает живое содержимое.
В делящейся клетке сначала образуется клеточная пластинка, превращающаяся в срединную пластинку, а затем формируется первичная оболочка. Она тонкая, эластичная, состоит в основном из пектиновых веществ (пектатов кальция, магния), целлюлозы и гемицеллюлоз. По мере роста и специализации определенных клеток их оболочка утолщается путем наслоения и образования вторичной оболочки. Она может оставаться целлюлозной эластичной или претерпевать химические изменения, терять эластичность, приобретать дополнительные свойства. В результате клеточная стенка слагается из первичной и вторичной оболочек. Опорно-структурными единицами вторичной клеточной оболочки являются молекулы целлюлозы, объединенные в цепочки — мицеллы. Пучки мицелл образуют микрофибриллы, собранные в волокнистые фибриллы.
Целлюлоза, или клетчатка, является скелетным веществом клеточной оболочки высших растений. Она образует структурный остов, в который погружены вещества двух других типов. Поэтому удаление этих веществ не вызывает значительного изменения формы оболочки и снижения ее механических свойств. Полимерная молекула целлюлозы состоит из большого числа мономеров — остатков глюкозы, связанных между собой в виде цепочки. Число этих остатков в молекуле целлюлозы неодинаково в клеточных оболочках разных растений, что и определяет различные свойства целлюлозы у разных видов. Целлюлоза обладает двойным лучепреломлением, характерным для кристаллов, однако она не типичный кристалл, так как способна к набуханию, а двойное лучепреломление объясняется строгой трехмерной пространственной ориентацией ее молекул, располагающихся параллельно друг другу. Целлюлоза имеет большое промышленное значение, так как все текстильные товары растительного происхождения и бумага состоят из целлюлозы, полученной из клеточных оболочек растений.
Гемицеллюлозы, иногда называемые также полуклетчатками, представляют собой группу полимерных углеводов, в основе которых лежит не глюкоза, как у целлюлозы, а другие мономеры— различные сахара, важнейшими из которых являются ксилоза, манноза и изомер глюкозы, в меньшей степени — арабиноза и галактоза. В состав одной молекулы гемицеллюлозы могут входить остатки сахара не одного типа, а нескольких, например, арабиноза в сочетании с ксилозой, изомер глюкозы в сочетании с маннозой (смешанные гемицеллюлозы). Называются гемицеллюлозы по составу образующих их мономеров с прибавлением окончания «ан». Например, гемицеллюлоза ксилан состоит из остатков ксилозы, маннан — из остатков маннозы, а в состав наиболее важной Гемицеллюлозы древесины хвойных — глюкоманнана — входят остатки глюкозы и маннозы.
Как и для целлюлозы, для гемицеллюлоз характерно нитчатое строение молекул, однако эти молекулы не имеют строгой пространственной ориентации, но и не являются аморфными. Подобно целлюлозе, они выполняют в теле растения механическую функцию, но некоторые из них могут выполнять также функцию запаса, отлагаясь и вновь расходуясь по мере потребности растения (например, в семенах и плодах).
Пектиновые вещества представляют собой продукты полимеризации близкой к углеводам галактуроновой кислоты (полиурониды), а также сахаров арабинозы (арабинаны) и галактозы (галактаны). Как и у гемицеллюлоз, в состав их молекул могут входить несколько мономеров в различных сочетаниях. Пектиновые вещества отличаются от гемицеллюлоз тем, что образуют не нитчатые, а сильно разветвленные молекулы, без всякой пространственной ориентации и поэтому встречаются в клетке, как правило, в аморфном состоянии. Второй особенностью пектиновых веществ является их высокая гидрофильность, набухаемость в воде и способность образовывать гели и слизи. Благодаря присутствию карбоксильных групп (СООН), они способны образовывать не растворимые в воде соли (пектаты), чаще всего с кальцием и магнием. Пектиновые вещества связаны с гемицеллюлозами постепенными, переходами.
В живых клетках оболочка всегда пропитана водой и находится в набухшем состоянии. Особенно много воды (до 80—90%) содержат оболочки молодых клеток. Оболочка более взрослых и особенно стареющих клеток может инкрустироваться различными веществами, например, лигнином, вызывающим ее одревеснение.
Состав веществ, принимающих участие в образовании клеточной оболочки, зависит таким образом от возраста клетки, а также от вида растения, органа и ткани, в которых данная клетка расположена. Клеточная стенка бесцветна и прозрачна, легко пропускает солнечный свет. Обычно стенки пропитаны водой. По системе клеточных стенок осуществляется транспорт воды и растворенных в ней низкомолекулярных соединений.
Клеточные стенки растений часто подвергаются химическим видоизменениям. Одревеснение, или лигнификацияпроисходит в том случае, если в матриксе откладывается лигнин – полимерное соединение фенольной природы, нерастворимое в воде. Одревесневшая клеточная стенка теряет эластичность, резко повышается ее твердость и прочность на сжатие, снижается проницаемость для воды. Реактивами на лигнин являются: 1) флороглюцин и концентрированная хлористоводородная или серная кислота (одревесневшие стенки приобретают вишнево-красную окраску) и 2) сульфат анилина, под действием которого одревесневшие стенки становятся лимонно-желтыми. Лигнификация характерна для стенок клеток проводящей ткани ксилемы (древесины) и механической ткани склеренхимы.
Опробковение, или суберинизация происходит в результате отложения с внутренней стороны клеточной стенки гидрофобных полимеров – суберина и воска. Суберин представляет собой смесь эфиров полимерных жирных кислот. Мономерами воска являются жирные спирты и восковые эфиры. Воск легко извлекается органическими растворителями и быстро плавится, образует кристаллы. Суберин – аморфное соединение, не плавится и не растворяется в органических растворителях. Суберин и воск, образуя чередующиеся параллельные слои, выстилают всю полость клетки с внутренней стороны в виде пленки. Субериновая пленка практически непроницаема для воды и для газов, поэтому после ее образования клетка обычно отмирает. Опробковение характерно для стенок клеток покровной ткани пробки. Реактивом на опробковевшую клеточную стенку является судан III, окраска оранжево-красная.
Кутинизации подвергаются наружные стенки клеток покровной ткани эпидермы. Кутин и воск откладываются чередующимися слоями на наружной поверхности клеточной стенки в виде пленки – кутикулы. Кутин представляет собой жироподобное полимерное соединение, близкое по химической природе и свойствам суберину. Кутикула предохраняет растение от излишнего испарения воды с поверхности растения. Окрасить ее можно реактивом судан III в оранжево-красный цвет.
Минерализацияклеточной стенки происходит вследствие отложения в матриксе большого количества минеральных веществ, чаще всего кремнезема (оксида кремния), реже оксалата и карбоната кальция. Минеральные вещества придают стенке твердость и хрупкость. Отложение кремнезема характерно для клеток эпидермы хвощей, осок и злаков. Приобретенная в результате окремнения жесткость стеблей и листьев служит защитным средством против улиток, а также значительно снижает поедаемость и кормовую ценность растений.
У некоторых специализированных клеток наблюдается ослизнение клеточной стенки. При этом вместо целлюлозной вторичной стенки происходит отложение аморфных, сильно гидратированных кислых полисахаридов в виде слизей и камедей, близких по химической природе к пектиновым веществам. Слизи хорошо растворяются в воде с образованием слизистых растворов. Камеди клейкие, вытягиваются в нити. В сухом виде они имеют роговую консистенцию. При отложении слизи протопласт постепенно оттесняется к центру клетки, его объем и объем вакуоли постепенно уменьшаются. В конце концов, полость клетки может целиком заполниться слизью, и клетка отмирает. В некоторых случаях слизь может проходить через первичную клеточную стенку на поверхность. В синтезе и секреции слизи основное участие принимает аппарат Гольджи.
Выделяемая растительными клетками слизь выполняет различные функции. Так, слизь корневого чехлика служит в качестве смазки, облегчающей рост кончика корня в почве. Слизевые железки насекомоядных растений (росянка) выделяют ловчую слизь, к которой приклеиваются насекомые. Слизь, выделяемая наружными клетками семенной кожуры (лен, айва, подорожники), закрепляет семя на поверхности почвы и защищает проросток от высыхания. Слизь окрашивается реактивом метиленовый синий в голубой цвет. Выделение камедей обычно происходит при поранении растений. Например, камедетечение из пораненных участков стволов и ветвей часто наблюдается у вишни и сливы. Вишневый клей представляет собой застывшую камедь. Камедь выполняет защитную функцию, закрывая рану с поверхности. Образуются камеди в основном у древесных растений из семейств бобовых (акации, трагакантовые астрагалы) и розоцветных подсемейства сливовых (вишня, слива, абрикос). Камеди и слизи используются в медицине.
Образование клеточной стенки начинается во время деления клетки. В плоскости деления образуется клеточная пластинка, единый слой, общий для двух дочерних клеток. Она состоит из пектиновых веществ, имеющих полужидкую консистенцию; целлюлоза отсутствует. Во взрослой клетке клеточная пластинка сохраняется, но претерпевает изменения, поэтому ее называют срединной, или межклеточной пластинкой.
Стенки делящихся и растущих клеток называют первичными. Они содержат много (60-90%) воды. В сухом веществе преобладают матричные полисахариды (60-70%), содержание целлюлозы не превышает 30%, лигнин отсутствует. Толщина первичной стенки очень невелика (0,1-0,5 мкм).Для многих клеток отложение клеточной стенки прекращается одновременно с прекращением роста клетки. Такие клетки окружены тонкой первичной стенкой до конца жизни.
У других клеток отложение стенки продолжается и по достижении клеткой окончательного размера. При этом толщина стенки увеличивается, а объем, занимаемый полостью клетки, сокращается. Такой процесс носит название вторичного утолщения стенки, а саму стенку называют вторичной. Вторичная стенка может рассматриваться как дополнительная, выполняющая главным образом механическую, опорную функцию. Именно вторичная стенка ответственна за свойства древесины, текстильного волокна, бумаги. Вторичная стенка содержит значительно меньше воды, чем первичная; в ней преобладают микрофибриллы целлюлозы (40-50% от массы сухого вещества), которые располагаются параллельно друг другу. Из полисахаридов матрикса характерны гемицеллюлозы (20-30%), пектиновых веществ очень мало. Вторичные клеточные стенки, как правило, подвергаются одревеснению. В неодревесневших вторичных стенках (лубяные волокна льна, волоски хлопчатника) содержание целлюлозы может достигать 95%. Большое содержание и строго упорядоченная ориентация микрофибрилл определяют высокие механические свойства вторичных стенок. Часто клетки, имеющие вторичную одревесневшую клеточную стенку, после завершения вторичного утолщения отмирают.
Срединная пластинка склеивает соседние клетки. Если ее растворить, стенки клеток теряют связь друг с другом и разъединяются. Этот процесс называется мацерация. Довольно обычна естественная мацерация, при которой пектиновые вещества срединной пластинки переводятся в растворимое состояние с помощью фермента пектиназы и затем вымываются водой (перезрелые плоды груши, дыни, персика, банана). Часто наблюдается частичная мацерация, при которой срединная пластинка растворяется не по всей поверхности, а лишь в углах клеток. Вследствие тургорного давления соседние клетки в этих местах округляются, в результате чего образуются межклетники .Межклетники образуют единую разветвленную сеть, которая заполняется парами воды и газами. Таким образом, межклетники улучшают газообмен клеток.
Характерная особенность вторичной стенки – ее неравномерное отложение поверх первичной стенки, в результате чего во вторичной стенке остаются неутолщенные участки – поры. Если вторичная стенка не достигает большой толщины, поры выглядят как мелкие углубления. У клеток с мощной вторичной стенкой поры в разрезе имеют вид радиальных каналов, идущих от полости клетки до первичной стенки. По форме порового канала различают поры двух типов – простые и окаймленные.
У простых пор диаметр порового канала по всей длине одинаковый и имеет форму узкого цилиндра. Простые поры характерны для паренхимных клеток, лубяных и древесинных волокон. У окаймленных порканал резко суживается в процессе отложения клеточной стенки, поэтому внутреннее отверстие поры, выходящее в полость клетки, гораздо уже, чем наружное, упирающееся в первичную стенку. Окаймленные поры характерны для рано отмирающих клеток водопроводящих элементов древесины. У них поровый канал по направлению к замыкающей пленке воронковидно расширяется, а вторичная стенка нависает в виде валика над расширенной частью канала, образуя камеру поры.
Поры облегчают транспорт воды и растворенных веществ от клетки к клетке, в то же время, не снижая прочности клеточной стенки.
3.Меристиматические и покровные ткани.
Ткани – этоустойчивые, т.е. закономерно повторяющиеся комплексы клеток, сходные по происхождению, строению и приспособленные к выполнению одной или нескольких функций.
Ткани возникли у растений в связи с переходом их предков – водорослей к наземному образу жизни. Ткани классифицируют в зависимости от выполняемой функции. Однако надо помнить, что одна и та же ткань может выполнять несколько функций. Различают сложные ткани, состоящие из разнородных элементов, выполняющих разные функции, и простые ткани, состоящие из однородных элементов. Функции ткани могут меняться в зависимости от ее возраста. Наиболее часто выделяют шесть типов тканей: 1) образовательные, или меристемы; 2) основные; 3) покровные; 4) выделительные; 5) механические; 6) проводящие.
Тело растения формируется в результате деятельности образовательных тканей, называемых меристемами. Основное свойство меристем – способность к делению и образованию новых клеток. В теле растения образовательные ткани функционируют в течение всей жизни. У векового дерева, наряду с очень старыми тканями можно обнаружить и молодые.
В составе меристем различают: 1) инициальные клетки, или инициали, и 2) производные от инициалей. Инициали сохраняют способность к делению в течение всей жизни растения и всегда остаются в составе меристем. Производные от инициалей делятся некоторое число раз и затем превращаются в постоянные ткани.
По происхождению различают: 1) первичные меристемы, которые берут начало непосредственно от меристем зародыша, и 2) вторичные меристемы, образующиеся на более поздних этапах развития растения либо из первичных меристем, либо в результате дедифференциации постоянных тканей. Постоянные ткани, образовавшиеся из первичных меристем, называются первичными, из них складывается первичная структура тела растения. Из вторичных меристем образуются вторичные ткани, которые определяют вторичный рост растения.
В зависимости от местоположения выделяют четыре типа меристем: 1) верхушечные, или апикальные; 2) боковые, или латеральные; 3) вставочные, или интеркалярные; 4) раневые, или травматические.
Верхушечные (апикальные) меристемы закладываются с первых стадий развития зародыша на верхушке побега и на кончике зародышевого корешка. По мере роста и ветвления на каждом боковом побеге и каждом корне образуются свои верхушечные (апикальные) меристемы. Они обеспечивают рост этих органов в длину. Апикальные меристемы всегда первичны, они образуют конусы нарастания корня и побега.
Боковые (латеральные) меристемы располагаются по окружности осевых органов (корней, стеблей) в виде цилиндров, которые на поперечных срезах имеют вид колец. Первичные боковые меристемы – прокамбий, перицикл – возникают непосредственно под апексами и в непосредственной связи с ними. Вторичные латеральные меристемы: камбий – возникает из прокамбия - и пробковый камбий (феллоген) - образуется из клеток постоянных тканей. Боковые меристемы обеспечивают рост корня и стебля в толщину. Из прокамбия и камбия образуются проводящие ткани, из феллогена – перидерма.
Вставочные (интеркалярные) меристемы находятся в основаниях междоузлий побегов и молодых листьев. Они первичны, поскольку являются остатками верхушечных меристем, их дифференциация задерживается по сравнению с остальными тканями. Вставочные меристемы не имеют в своем составе инициалей и со временем полностью превращаются в постоянные ткани. В их составе могут присутствовать некоторые дифференцированные элементы, например проводящие. Интеркалярный рост характерен для стеблей злаков, наблюдается также в основании луковиц, завязей.
Раневые ( травматические) меристемы обычно образуются при повреждении тканей и органов. Живые клетки постоянных тканей, окружающие пораженные участки, дедифференцируются и начинают делиться, т.е. превращаются во вторичную меристему. Раневые меристемы образуют каллус – особую ткань, состоящую из однородных паренхимных клеток, прикрывающих место поранения. Из каллуса может возникнуть любая ткань или орган растения. Часто клетки формируют феллоген, образующий на поверхности перидерму, которая закрывает рану и способствует ее заживлению. Способность растений к каллусообразованию используют для получения культуры изолированных тканей, а также в практике садоводства для размножения растений черенками и прививками.
Типичные цитологические признаки образовательных тканей наиболее отчетливо выражены у апикальных меристем. Это изодиаметрические многогранные клетки, не разделенные межклетниками. Клеточные стенки тонкие, с малым содержанием целлюлозы. Цитоплазма густая, ядро относительно крупное, занимает центральное положение. В цитоплазме большое число рибосом и митохондрий, так как происходит интенсивный синтез белков и других веществ. Вакуоли очень мелкие, многочисленные.
Покровные ткани располагаются на поверхности органов растений на границе с внешней средой. Они состоят из плотно сомкнутых клеток и защищают внутренние части растения от неблагоприятных внешних воздействий, излишнего испарения и иссушения, резкой перемены температуры, проникновения микроорганизмов, служат для газообмена и транспирации. В соответствии с происхождением из различных меристем выделяют первичные и вторичные покровные ткани.
К первичным покровным тканям относят: 1) ризодерму, или эпиблему и 2) эпидерму.
Ризодерма (эпиблема) – первичная однослойная поверхностная ткань корня. Образуется из протодермы – наружного слоя клеток апикальной меристемы корня. Основная функция ризодермы – всасывание, избирательное поглощение из почвы воды с растворенными в ней элементами минерального питания. Через ризодерму происходит выделение веществ, действующих на субстрат и преобразующих его. Клетки ризодермы тонкостенные, с вязкой цитоплазмой и большим количеством митохондрий (минеральные ионы поглощаются активно, с затратой энергии, против градиента концентрации). Характерной особенностью ризодермы является образование у части клеток корневых волосков – трубчатых выростов, в отличие от трихомов не отделенных стенкой от материнской клетки Корневые волоски увеличивают поглощающую поверхность ризодермы в десять и более раз. Волоски имеют длину 1-2 (3) мм. Ризодерму часто рассматривают как всасывающую ткань.
Эпидерма - первичная покровная ткань, образующаяся из протодермы конуса нарастания побега. Она покрывает листья, стебли травянистых и молодых побегов древесных растений, цветки, плоды и семена. Основная функция эпидермы – регуляция газообмена и транспирации (испарения воды живыми тканями). Кроме того, эпидерма выполняет целый ряд других функций. Она препятствует проникновению внутрь растения болезнетворных организмов, защищает внутренние ткани от механических повреждений и придает органам прочность. Через эпидерму могут выделяться наружу эфирные масла, вода, соли. Эпидерма может функционировать как всасывающая ткань. Она принимает участие в синтезе различных веществ, в восприятии раздражений, в движении листьев.
Эпидерма - сложная ткань, в ее состав входят морфологически различные типы клеток: 1) основные клетки эпидермы; 2) замыкающие и побочные клетки устьиц; 3) трихомы.
Основные клетки эпидермы – живые клетки таблитчатой формы. Вид клеток с поверхности различен. Клетки плотно сомкнуты, межклетники отсутствуют. Боковые стенки (перпендикулярные поверхности органа) часто извилистые, что повышает прочность их сцепления, реже прямые. Эпидермальные клетки осевых органов и листьев многих однодольных сильно вытянуты вдоль оси органа.
Наружные стенки клеток обычно толще остальных. Их внутренний, более мощный, слой состоит из целлюлозы и пектиновых веществ; наружный слой подвергается кутинизации. Поверх наружных стенок выделяется сплошной слой кутина, образующий защитную пленку – кутикулу. Помимо кутина в ее состав входят вкрапления воска, что еще больше снижает проницаемость кутикулы для воды и для газов. Воск может откладываться в кристаллической форме и на поверхности кутикулы в виде чешуек, палочек, трубочек и других структур, видимых только в электронный микроскоп. Этот сизый, легко стирающийся налет хорошо заметен на листьях капусты, плодах сливы, винограда. Мощность кутикулы, распределение в ней восков и кутина определяют химическую стойкость и проницаемость эпидермы для газов и растворов. В условиях засушливого климата у растений развивается более толстая кутикула. У растений, погруженных в воду, кутикула отсутствует.
Клетки эпидермы имеют живой протопласт, обычно с хорошо развитой эндоплазматической сетью и аппаратом Гольджи. У большинства видов растений в цитоплазме присутствуют лейкопласты. У водных растений, папоротников, обитателей тенистых мест (гибискус) встречаются редкие хлоропласты. Эпидерма чаще всего состоит из одного слоя клеток. Редко встречается двух- или многослойная эпидерма, преимущественно у тропических растений, живущих в условиях непостоянной обеспеченности водой (бегонии, пеперомии, фикусы). Нижние слои многослойной эпидермы функционируют как водозапасающая ткань. У некоторых растений клеточные стенки могут пропитываться кремнеземом (хвощи, злаки, осоки) или содержать слизи (семена льна, айвы, подорожников).
Устьица – образования для регуляции транспирации и газообмена. Устьице состоит из двух замыкающих клеток бобовидной формы, между которыми находится устьичная щель, которая может расширяться и сужаться. Под щелью располагается крупный межклетник – подустьичная полость. Клетки эпидермы, примыкающие к замыкающим клеткам, часто отличаются от остальных клеток, и тогда их называют побочными, или околоустьичными клетками. Они участвуют в движении замыкающих клеток.
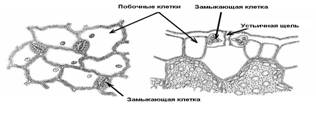
Схема строения устьица.
Замыкающие и побочные клетки образуют устьичный аппарат. В зависимости от числа побочных клеток и их расположения относительно устьичной щели выделяют несколько типов устьичного аппарата.
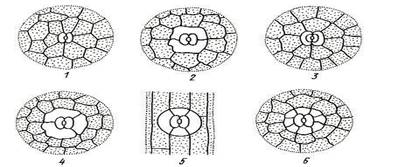
Типы устьичного аппарата : 1 – аномоцитный; 2 – диацитный; 3 – парацитный; 4 – анизоцитный; 5 – тетрацитный; 5 – энциклоцитный.
Аномоцитный тип устьичного аппарата обычен для всех групп растений, исключая хвощи. Побочные клетки в этом случае не отличаются от остальных клеток эпидермы.
Диацитный тип характеризуется двумя побочными клетками, которые располагаются перпендикулярно устьичной щели. Этот тип обнаружен у некоторых цветковых растений, в частности, у большинства губоцветных (мята, шалфей, чабрец, душица) и гвоздичных. При парацитном типе две побочные клетки располагаются параллельно замыкающим и устьичной щели. Он найден у папоротников, хвощей и ряда цветковых растений.
Анизоцитный тип обнаружен только у цветковых растений, в частности, он встречается у крестоцветных (пастушья сумка, желтушник) и пасленовых (белена, дурман, красавка). В этом случае замыкающие клетки окружены тремя побочными, одна из которых заметно крупнее или мельче остальных.
Тетрацитным типом устьичного аппарата характеризуются преимущественно однодольные.
При энциклоцитном типе побочные клетки образуют узкое кольцо вокруг замыкающих клеток. Подобная структура найдена у папоротников, голосеменных и некоторых цветковых.
У многих растений эпидерма образует наружные одно- или многоклеточные выросты различной формы – трихомы.
Трихомы делятся на: 1) кроющие и 2) железистые.
Кроющие трихомы имеют вид простых, разветвленных или звездчатых волосков, одно- или многоклеточных. Кроющие трихомы могут длительное время оставаться живыми, но чаще они быстро отмирают и заполняются воздухом.
Густой слой волосков отражает часть солнечных лучей и уменьшает нагрев, создает затишное пространство около эпидермы, что в совокупности снижает транспирацию. Часто волоски образуют покров только там, где располагаются устьица, например на нижней стороне листьев мать-и-мачехи, багульника. Жесткие, колючие волоски защищают растения от поедания животными, сосочки на лепестках привлекают насекомых.
К вторичным покровным тканям относятся: 1) перидерма и 2) корка, или ритидом.
Перидерма – сложная многослойная покровная ткань, которая приходит на смену первичным покровным тканям – ризодерме и эпидерме. Перидерма покрывает корни вторичного строения и стебли многолетних побегов. Она может возникнуть и в результате залечивания поврежденных тканей раневой меристемой. Перидерма состоит из трех комплексов клеток, различных по строению и функциям. Это: 1) феллема, или пробка, выполняющая главные защитные функции; 2) феллоген, или пробковый камбий, за счет работы которого образуется перидерма в целом; 3) феллодерма, или пробковая паренхима, выполняющая функцию питания феллогена.
Главная функция пробки – защита от потери влаги. Кроме того, пробка предохраняет растение от проникновения болезнетворных организмов, а также дает механическую защиту стволам и ветвям деревьев, а феллоген залечивает нанесенные повреждения, образуя новые слои пробки. Поскольку клетки пробки заполнены воздухом, пробковый футляр обладает малой теплопроводностью и хорошо предохраняет от резких колебаний температуры.
У большинства деревьев и кустарников феллоген закладывается в однолетних побегах уже в середине лета. Чаще всего он возникает из паренхимных клеток, лежащих сразу под эпидермой. Иногда феллоген образуется в более глубоких слоях коры (смородина, малина). Редко эпидермальные клетки, делясь, превращаются в феллоген (ива, айва, олеандр).
Газообмен и транспирация в органах, покрытых перидермой, происходят через чечевички. В местах чечевичек пробковые слои разорваны и чередуются с паренхимными клетками, рыхло соединенными между собой. По межклетникам этой выполняющей ткани циркулируют газы. Феллоген подстилает выполняющую ткань и, по мере ее отмирания, дополняет новыми слоями. С наступлением холодного сезона феллоген откладывает под выполняющей тканью замыкающий слой, состоящий из клеток пробки. Весной этот слой под напором новых клеток разрывается. В замыкающих слоях имеются небольшие межклетники, так что живые ткани ветвей деревьев даже зимой не отграничены наглухо от окружающей среды.
У большинства древесных растений на смену гладкой перидерме приходит трещиноватая корка (ритидом) . У сосны это происходит на 8-10-м году, у дуба – в 25-30 лет, у граба – в 50 лет. Лишь у некоторых деревьев (осина, бук, платан, эвкалипт) корка вообще не образуется.
Корка возникает в результате многократного заложения новых прослоек перидермы во все более глубоких слоях коры. Живые клетки, заключенные между этими прослойками, погибают. Таким образом, корка состоит из чередующихся слоев пробки и прочих отмерших тканей коры. Мертвые ткани корки не могут растягиваться, следуя за утолщением ствола, поэтому на стволе появляются трещины, не доходящие, однако, до глубинных живых тканей. Граница между перидермой и коркой внешне заметна по появлению этих трещин, особенно ясна эта граница у березы, у которой белая береста (перидерма) сменяется черной трещиноватой коркой. Толстая корка надежно предохраняет стволы деревьев от механических повреждений, лесных пожаров, резкой смены температур.
4.Механические и проводящие ткани.
Механические (скелетные, опорные, арматурные) ткани выполняют в растении роль скелета, который скрепляет ткани и части органов между собой. Они придают растениям прочность, способность противостоять действию тяжести собственных органов, порывам ветра, дождю, снегу, вытаптыванию животными. Клетки механических тканей разнообразны по форме, но имеют общий признак – сильно утолщенные клеточные стенки, которые даже после отмирания протопласта продолжают выполнять опорную функцию. Различают два типа механических тканей: 1) колленхиму и 2) склеренхиму.
Колленхима – механическая ткань молодых растущих органов, возникает очень рано, когда еще продолжается рост органа в длину. Колленхима состоит из живых, вытянутых по оси органа клеток с тупыми или скошенными концами. В клетках часто содержатся хлоропласты. Клеточные стенки утолщены неравномерно и никогда не одревесневают. Граница между первичной и вторичной стенками не выражена, в утолщениях чередуются слои целлюлозы и сильно обводненные слои, богатые пектинами и гемицеллюлозами. В растущем органе стенки клеток должны сохранять способность к растяжению, что возможно только в живых клетках. Функции опорной ткани колленхима может выполнять только в состоянии тургора. Если растение теряет воду, тонкие участки клеточных стенок складываются «гармошкой», побеги теряют упругость и обвисают.
Колленхима располагается сразу за покровной тканью в молодых стеблях, цветоносах, черешках листьев, образуя сплошной цилиндр или тяжи в ребрах. В корнях колленхима обычно не встречается. Наиболее характерна колленхима для двудольных, у однодольных растений она встречается редко.
В зависимости от характера утолщения стенок клеток различают три типа колленхимы: 1) уголковую, 2) пластинчатую и 3) рыхлую.
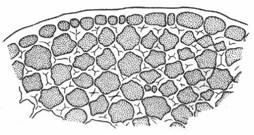
Уголковая колленхима на поперечном срезе листового черешка свеклы.
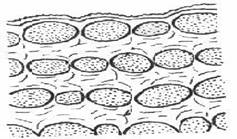
Пластинчатая колленхима на поперечном срезе.
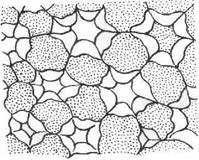
Рыхлая колленхима на поперечном срезе.
Склеренхима встречается наиболее часто, во всех органах: корнях, стеблях, листьях, плодах, цветках, семенах. Клетки склеренхимы имеют равномерно утолщенные и, как правило, одревесневшие стенки. Полость клетки мала, поры простые, щелевидные, немногочисленные. Протопласт, как правило, рано отмирает, и опорную функцию выполняют мертвые клетки. Различают два типа склеренхимы: 1) волокна и 2) склереиды, различающиеся формой клеток.
Волокна – прозенхимные клетки, сильно вытянутые в длину и заостренные на концах. Они обеспечивают прочность органов растений на растяжение, сжатие и изгибы. Прочность волокон повышается благодаря тому, что фибриллы целлюлозы проходят в них винтообразно, меняя направление во внешних и внутренних витках.
Склереиды – клетки, имеющие различную форму, чаще паренхимную. Они встречаются как поодиночке, в виде идиобластов, так и группами. Это мертвые клетки с очень толстыми одревесневшими стенками, пронизанными поровыми каналами, которые часто ветвятся В зависимости от формы клеток выделяют несколько типов склереид. Наиболее часто встречаются брахисклереиды, или каменистые клетки и астросклереиды.

Каменистые клетки имеют более или менее округлую форму.Из них состоят косточки вишни, сливы, персика, скорлупа грецкого ореха. Они встречаются в мякоти плодов груши, айвы, рябины, в корнях хрена среди тонкостенных клеток. У груши при созревании плода наблюдается раздревеснение каменистых клеток.
Астросклереиды имеют ветвистую форму с отростками, направленными в разные стороны
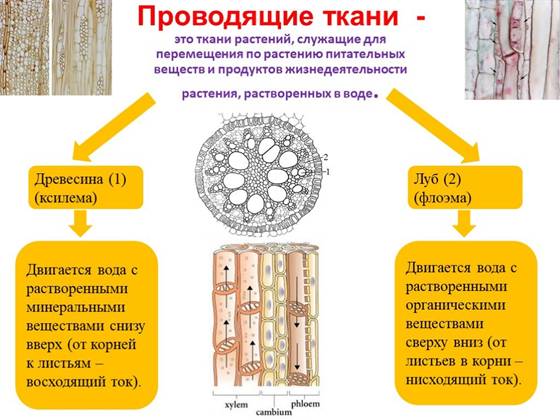
Проводящие ткани служат для передвижения по растению растворенных в воде питательных веществ. Они возникли как следствие приспособления растений к жизни на суше. В связи с жизнью в двух средах – почвенной и воздушной, возникли две проводящие ткани, по которым вещества передвигаются в двух направлениях. По ксилеме от корней к листьям поднимаются вещества почвенного питания – вода и растворенные в ней минеральные соли (восходящий, или транспирационный ток). По флоэме от листьев к корням передвигаются вещества, образовавшиеся в процессе фотосинтеза, главным образом сахароза (нисходящий ток). Так как эти вещества представляют собой продукты ассимиляции углекислого газа, транспорт веще ств по флоэме называют током ассимилятов.
Проводящие ткани образуют в теле растения непрерывную разветвленную систему, соединяющую все органы – от тончайших корешков до самых молодых побегов. Ксилема и флоэма представляют собой сложные ткани, в их состав входят разнородные элементы – проводящие, механические, запасающие, выделительные. Самыми важными являются проводящие элементы, именно они выполняют функцию проведения веществ.
Ксилема и флоэма формируются из одной и той же меристемы и, поэтому, в растении всегда располагаются рядом. Первичные проводящие ткани образуются из первичной латеральной меристемы – прокамбия, вторичные – из вторичной латеральной меристемы – камбия. Вторичные проводящие ткани имеют более сложное строение, чем первичные.
5.Строение пучкового и беспучкового стебля.
Стебель представляет собой ось побега, состоящую из узлов и междоузлий. Основные функции стебля – опорная (несущая) и проводящая. Стебель осуществляет связь между корнями и листьями. В многолетних стеблях обычно откладываются запасные питательные вещества. Молодые стебли, имеющие под эпидермой хлоренхиму, активно участвуют в фотосинтезе. Стебель обычно имеет цилиндрическую форму и характеризуется радиальной симметрией в расположении тканей.
Различают два основных типа стеблей: травянистый и деревянистый. Травянистый существует обычно один вегетационный сезон и отличается слабым утолщением; деревянистый обычно многолетний, утолщающийся неопределенно долго. Стебли картофеля, томатов, злаков в поперечном сечении округлые; стебель тыквы — многогранный, шалфея — четырехгранный, а осок — трехгранный.
По форме роста и расположению в пространстве различают прямостоячие стебли (травянистый подсолнечника, деревянистый — ствол березы, дуба); ползучие (гусиная лапка, земляника), их называют усами; вьющиеся (вьюнок полевой, хмель); лазающие, которые прикрепляются к опоре при помощи усиков (горох, вика).
У некоторых растений стебель недоразвивается и листья у них образуют прикорневую розетку (одуванчик, подорожник, свекла, морковь).
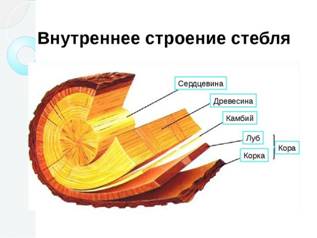
Сердцевиназанимает центральную часть цилиндра, паренхимные клетки её вытянута по оси стебля, к самому центру уменьшаются. Нередко они разрыхляются отмирают и заполняются воздухом (подсолнечник, кукуруза, бузина). Если отмирание сердцевины происходит раньше, чем заканчивается рост стебля, то происходит разрыв сердцевины и образование полого стебля (тыквенные, зонтичные, злаки).
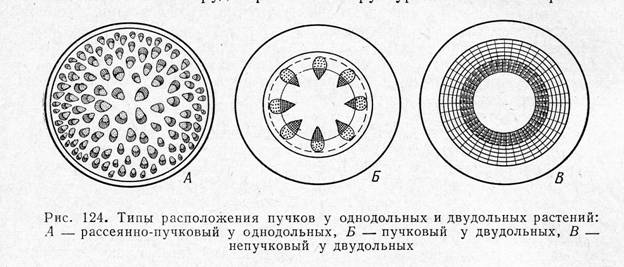
1)Непучковый цилиндр - кольца первичной флоэмы и ксилемы образуют более или менее сплошной полый цилиндр первичных проводящих тканей, погруженных в основную ткань. Снаружи от цилиндра формируется первичная кора, а внутри – сердцевина.(древесные растения, тысячелистник, лен)
2)Пучковый цилиндр – система отдельных проводящих пучков, образованных первичной ксилемой и флоэмой; располагаются по кругу кольцом и разделены первичными сердцевинными лучами(травянистые двудольные).
Камбий: 1)Пучковый – это камбий, имеющийся в сосудистых пучках. Преобразуются в элементы вторичной флоэмы и ксилемы.
2) Межпучковый(лучевой) – это камбий, находящийся между пучками в сердцевинных лучах; преобразуется только в паренхиму, образующую сердцевинные лучи
Строение стебля:
1. Пучковый – тип строения стебля при котором проводящие комплексы – флоэма и ксилема входят в состав проводящих пучков, располагаются либо по кругу(у двудольных), либо по всей толще стебля(у однодольных)
2. Непучковый- тип строения стебля, при котором флоэма и ксилема располагаются в нем сплошными цилиндрами: ксилема-ближе к центру, флоэма к периферии от ксилемы. Между ними находиться камбиальный цилиндр, производящий в центробежном направлении вторичную флоэму, а в центростемительном – вторичную ксилему.
6.Деятельность камбия.
Ткань, расположенная в дереве между древесиной и корой, является особой. Она состоит из живых клеток, способных к делению и росту. Такая ткань имеет название "камбий". Эта ткань образует вокруг внешней окружности древесины очень тонкое кольцо размером менее миллиметра, которое можно увидеть лишь с помощью увеличительных приборов. Клетки камбия имеют двойное назначение. При их делении вырабатываются как древесинные клетки, так и клетки коры. Свойства клеток, которые образует камбий, различны. Для древесной части создаются клетки, по которым движутся различные соки дерева и питательные вещества. Из них состоит проводящая ткань. При образовании коры создаются ткани механические, придающие крепость стволу.
Камбий представляет собой образовательный слой деятельных клеток, залегающий на границе между древесиной и лубом. Камбий происходит из прокамбия — родоначальной ткани сосудистых пучков, в свою очередь возникающей из клеток первичной образовательной ткани, первоткани, или иначе первичной меристемы. Сначала полоса камбия имеется только в сосудистых пучках (пучках открытых — они одни только имеют камбий). Это — камбий пучковый. Затем образуются прослойки камбия между пучками, в сердцевинных лучах (камбий межпучковый, или лучевой). Прослойки эти соединяют камбиальные полосы пучков друг с другом. Таким образом получается целое сплошное камбиальное кольцо, идущее параллельно окружности. Камбий межпучковый образует новые сосудистые пучки в промежутках между старыми, или же все кольцо однообразно функционирует, порождая ежегодно новые массы древесины и луба. В морозные зимы деятельность камбия полностью прекращается, а с нею и прирост дерева в толщину.
Камбиальные клетки обыкновенно вытянуты по длине, имеют вид прямоугольных 4-гранных призм. Внутри клетки находится густозернистая плазма и явственное веретенообразное ядро, вытянутое по направлению продольной оси клетки; иногда также хлорофилл, а зимою мелкие крупины крахмала. Оболочка клеток неодеревеневшая, нежная и тонкая, только на зиму утолщающая свои радиальные стенки. Клетки с только что описанными свойствами образуют несколько концентрических слоев, прилегающих друг к другу. За настоящий камбий принимают, однако, только один из этих слоев, остальные считают самыми молодыми древесиной и лубом. Таким образом, в каждом радиальном ряду клеток находится только одна настоящая камбиальная клетка, это так называемая инициальная клетка. Она делится продольно пополам тангентальной перегородкой на две клетки, из которых одна сохраняет свойства произведшей ее (материнской) клетки и прежде всего способность снова делиться — она становится новой инициальной клеткой, другая клетка делится еще раз (опять в тангентальной плоскости), превращаясь в пару клеток «постоянных», притом — лубяных, если они лежат к наружи от инициальной клетки, древесинных, если — внутри. Результатом повторных делений инициальной клетки является правильный радиальный ряд клеток древесины и луба. Клетки древесины отлагаются по направлению к центру стебля, клетки луба — к периферии. В древесине и в лубе наиболее молодые части лежат всего ближе к камбию; стало быть, у древесины самая молодая часть наружная, а у луба наоборот — внутренняя. Расширение самого камбиального кольца происходит благодаря делению клеток камбия радиальными перегородками.
7.Тканевый и клеточный состав древесины и луба.
Луб (флоэма) – это сложная проводящая ткань, по которой продукты фотосинтеза (органические вещества) транспортируются из листьев ко всем органам растения (к корневищам, плодам, семенам и т.д.). Образуется флоэма делением клеток прокамбия (первичная) и камбия (вторичная). Располагается луб в стебле снаружи от камбия под корой, а в листьях - ближе к нижней стороне пластинки. Под камбием в стволе находится древесина.
Ткани
флоэмы и ее клеточный состав делятся на три вида в зависимости от выполняемых
функций: ситовидные трубки с клетками; механические ткани (склереиды и
волокна); лубяная паренхима с паренхимными клетками. В основном, луб состоит из
ситовидных трубок, которые обеспечивают движение растворенных питательных веществ
вниз по стеблю. Трубки образуются ситовидными клетками, плотно прилегающими и
соединяющимися друг с другом.
Клетки
живые, тонкостенные и имеют удлиненную форму. У них отсутствует ядро, а в
центральной части содержится цитоплазма. Поперечные стенки клеток имеют мелкие
сквозные отверстия, через которые тяжи цитоплазмы переходят в соседние клетки.
Ситовидные трубки тянутся по всей длине растения. У лиственных растений к членикам ситовидных трубок примыкают и соединяются с ними клетки-спутники, которые также принимают участие в транспорте веществ. Функционируют ситовидные трубки недолго, лишь один вегетационный период, постепенно они закупориваются каллозой, а затем отмирают. Только у некоторых многолетних растений продолжительность жизни составляет более 2 лет.
Механические ткани – толстостенные лубяные волокна служат для прочности, а также выполняют опорную функцию. Лубяная паренхима содержит тонкостенные паренхимные клетки, которые служат для отложения запасных питательных веществ, а также их ближней транспортировки.
Если в ксилеме перемещение растворенных минеральных веществ осуществляется только вверх к листьям от корней, то в флоэме движение органических веществ (сахароза, углеводы, аминокислоты, фитогормоны) от листьев происходит к тем органам растения, которые их потребляют или запасают. Наибольшая интенсивность потребления веществ наблюдается в верхушках побегов, формирующихся листьях, корнях. Многие же растения имеют запасающиеся органы: клубни, луковицы и др. Скорость транспорта довольно большая и составляет десятки сантиметров в час. Экспериментами было установлено, что доноры-листья чаще всего подкармливают рядом расположенные органы растения. Например, листья побега обеспечивают плоды, нижние листья – корни. Кроме этого, транспорт флоэмы двусторонний, в зависимости от вегетационной фазы, например, запасающие органы могут транспортировать углеводы к распускающимся листьям.
Если кору на дереве по кругу перерезать до древесины, то органические вещества перестанут поступать к корням, и дерево со временем засохнет.
8.Годичные кольца древесины.
Каждый год при росте дерева в нем появляется наружный слой древесины в виде кольца. Такое образование называют годичным кольцом. Если сосчитать их число, то можно точно узнать возраст дерева. Однако, стоит отметить, что не у всех деревьев бывают годичные кольца, и не всегда они появляются в течении года.
Каждое годичное кольцо образуют две части: внутренняя часть кольца (светлая и рыхлая) и наружная часть (более темная и плотная). Эти две части одного и того же кольца различаются временем их образования: светлая и более рыхлая – весной и летом; темная—осенью. Подобный вид годичных слоев дерева характерен лишь для средней полосы. Зимой в нашем климате деревья не растут, поэтому они имеет неоднородную структуру.
Весной с наступлением тепла, когда прекращаются морозы и оттаивает земля, в дереве возобновляются жизненные процессы: начинается движение соков, рост цветочных почек, распускаются листья, появляются новые побеги, проходит цветение. Во время бурного весеннего развития дерево нуждается в ускоренной передаче воды и питательных веществ к ветвям из корней, которые, в свою очередь, впитывают необходимые элементы из почвы. На помощь приходит камбий. Он вырабатывает много клеток для построения проводящей ткани, которая состоит из широкопросветных сосудов. Через них необходимо будет пропустить большое количество необходимых соков. Нарождающаяся древесина создает внутреннюю, весеннюю часть годичного кольца.
К осени начинается процесс замедления роста и образуются узкопросветные сосуды с утолщенными стенками, которые придают прочность стволу. Полости клеток механической ткани значительно меньше, зато они имеют утолщенные стенки. Эти клетки создают наружную, осеннюю часть годичного кольца дерева, располагающуюся снаружи и уплотненную. На следующий год процесс возобновляется. Сперва образуются клетки проводящей ткани, а к концу года - механической.
Таким образом легко определить границу между древесиной двух смежных лет. Границей является линия соприкосновения клеток, отложенных осенью предыдущего года, с клетками, образовавшимися весной следующего года. У таких лиственных пород, как дуб и ясень, годичные образовния хорошо заметны, потому что в весенней части древесины у этих растений сосредоточено большое количество крупных сосудов. У других лиственных пород (березы, ивы, липы, осины) сосуды мелкие и годичный слой различим слабо. Для лабораторных исследований, чтобы граница между кольцами выступала резче, такую древесину нужно окрасить. Самый простой способ - использование химического карандаша. У хвойных древесных пород, например, ель, кедр, лиственница, сосна или пихта, годичные кольца всегда хорошо видны. У них поздняя осенняя древесина имеет окраску более темных тонов, чем весенняя.
Важно запомнить, что годичные кольца образуются из-за неравномерного деления клеток камбия!
9.Строение древесины хвойных деревьев. Смолоносная система.
К хвойным породам относятся: ель, лиственница, сосна,
кедр, пихта и др. 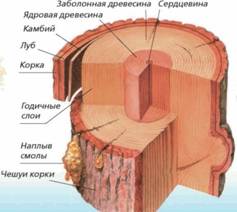
Древесина хвойных пород отличается сравнительно простым и правильным строением. Это следует из того, что она состоит всего лишь из двух видов клеток – трахеид и паренхимных клеток. Кроме того, анатомические элемента располагаются правильными радиальными рядами.
Трахеиды являются основным элементом древесины хвойных пород и занимают свыше 90% ее общего объема. Они имеют сильно вытянутую вдоль ствола веретенообразную форму с закругленными или кососрезанными концами. На поперечном срезе форма трахеид у большинства пород близка к прямоугольнику, а лиственницы – к пяти – или шестиугольнику.
Ранние трахеиды имеют сравнительно тонкие стенки и широкие полости, выполняют проводящую функцию. Поздние трахеиды имеют толстые стенки клеток и узкие полости, сплюснутые в радиальном направлении, выполняют механическую функцию, придавая древесине прочность. Вследствие различного строения поздних трахеид одного года и ранних последующего года у хвойных пород резко выражена граница годичных слоев.
Паренхимные клетки тонкостенные, с большой полостью, слегка вытянутые в радиальном направлении. В древесине хвойных пород паренхимные клетки в основном расположены в сердцевинных лучах, а также сопровождают смоляные ходы и служат для проведения и хранения запасных питательных веществ. Паренхимные клетки также выстилают каналы смоляных ходов и выполняют выделительную функцию. Кроме того, в древесине хвойных пород (кроме сосны и тисса) встречаются клетки, которые вытянуты по длине ствола и называются древесной паренхимой. Они расположены среди трахеид в виде одиночных клеток, а также сопровождают смоляные ходы (сопровождающая паренхима).
Сердцевинные лучи. В древесине всех хвойных пород сердцевинные лучи очень узкие (на поперечном разрезе однорядные, по высоте состоят из 3 – 5 рядов клеток). У сосны, кедра, лиственницы и ели сердцевинные лучи состоят из двух видов клеток: верхний и нижний ряды по высоте луча, состоят из горизонтальных (лучевых трахеид) с мелкими порами, средние по высоте – из паренхимных клеток с простыми порами. В сердцевинных лучах пихты, тисса, можжевельника лучевых трахеид нет.
Смоляные ходы. Смоляной ход – узкий, длинный межклеточный канал, образованный паренхимными клетками. Встречаются в древесине сосны, кедра, лиственницы, ели. Нет смоляных ходов у пихты, тисса, можжевельника. По расположению в стволе дерева выделяется два вида смоляных ходов: вертикальные и горизонтальные. Внутренний и внешний слой смоляного хода образованы живыми паренхимными клетками, средний – отмершими клетками, заполненными воздухом. Горизонтальные проходят по сердцевинным лучам и имеют два слоя клеток: эпителий (внутренний) и кольцо мертвых клеток. Диаметр горизонтальных смоляных ходов в 2,5 – 3 раза меньше диаметра вертикальных. Горизонтальные смоляные ходы пересекаются с вертикальными, образуя единую смолоносную систему.
При повреждении ствола растущего дерева образуются смоляные ходы, которые отличаются от нормальных, большим диаметром, неправильной формой, малой протяженностью вдоль ствола и образуют большие группы, вытянутые тангентальном направлении. Число клеток эпителия может достигать 30-ти (в нормальных смоляных ходах сосны – 4 – 6, ели – 6 – 9, лиственницы – до 12).
Патологические ходы могут появляться и в древесных породах, не имеющих нормальных смоляных ходов, например, в древесине пихты.
10.Строение и типы древесины лиственных деревьев. Возрастные изменения древесины.
Древесина лиственных пород, в отличие от хвойных, имеет более сложное строение: большее количество анатомических элементов, расположенных менее упорядоченно. Основные виды анатомических элементов: сосуды, волокна либриформа, паренхимные клетки. Переходные: сосудистые и волокнистые трахеиды, перегородочный либриформ.
В зависимости от особенностей размещены и величины, просветов сосудов в пределах годичного слоя древесину лиственных пород делят на кольцесосудистые и рассеянно-сосудистые.
Сосуды являются основными элементами для проведения в растущем дереве воды, с растворенными в ней, минеральными веществами, из корней к листьям. Сосуд – вертикальная трубка, состоящая из коротких клеток (члеников) с широкими полостями и тонкими стенками, длиною 2 см и более, достигая в отдельных случаях от 1,5 – 2 м (дуб). Перфорации (перегородки) между члениками сосудов могут быть простые у большинства пород и лестничные – береза, ольха, самшит, у бука и платана – оба вида перфораций.
Для части пород характерны утолщения стенок сосудов, которые придают им прочность. Спиральное утолщение сосудов имеется в древесине некоторых пород (граб. липа, вяз, клен и др.). У большинства пород сосуды лишены утолщений или они слабо выражены. Стенки сосудов имеют окаймленные поры, которые чаще образуют диагональные и реже – горизонтальные ряды. Окаймленные поры смежных сосудов отличаются от окаймленных пор хвойных пород меньшей величиной и отсутствием торусов. В местах примыкания к паренхимным клеткам поры обычно имеют окаймление со стороны сосудов (полуокаймленные поры). Между стенками сосудов и волокон либриформа поры отсутствуют. При образовании ядра сосуды закупориваются тиллами, представляющими собой мешковидные выросты соседних паренхимных клеток. Тиллы свойственны древесине ясеня, дуба, акации, ореха и некоторым другим породам.
Сосудистые трахеиды являются переходной формой между типичными трахеидами и сосудами. Они похожи на членики мелких сосудов по форме, размерам, порам и спиральным утолщениям. По сравнению с трахеидами хвойных пород сосудистые трахеиды имеют большую полость, слабо утолщенные оболочки и выполняют преимущественно проводящую функцию. Эти элементы встречаются лишь у некоторых пород (липа, осина, дуб и др.).
Волокна либриформа (от латинского «либри» – лыко и «форма» – вид) составляют 35 – 75% древесины лиственных пород и выполняют механическую функцию. Волокна либриформа сильно вытянутые вдоль оси ствола, клетки с заострёнными концами, толстыми стенками и узкими полостями. На поперечном срезе имеют округлую или многогранную форму. Поры на стенках древесных волокон немногочисленные, простые щелевидной формы и направлены под углом к продольной оси.
Наряду с типичным либриформом встречаются его разновидности: перегородочный либриформ, с поперечными перегородками и с менее тонкими стенками.
Волокнистая трахеида – переходная форма от типичной трахеиды к волокну либриформа. В отличие от волокон либриформа они имеют более заметную полость, меньшую толщину стенок и окаймленные поры. Волокнистые трахеиды в растущем дереве выполняют как механическую, так и проводящую функции. Они составляют значительную часть массы древесины березы, ольхи, граба и других пород.
Паренхимные клетки образуют в стволе дерева две системы: горизонтальную (сердцевинные лучи) и вертикальную (древесная паренхима). В растущем дереве служат для хранения питательных веществ. В древесине лиственных пород паренхимные клетки, составляют 8 – 40% объема древесины ствола.
Сердцевинные лучи состоят только из паренхимных клеток. Сердцевинные лучи у лиственных пород занимают значительно больший объем ствола, чем у хвойных. По ширине сердцевинные лучи делятся на однорядные, многорядные и ложноширокие. Однорядные имеют по ширине один ряд клеток (осина, ива, тополь, береза.). Многорядные лучи заключают по ширине два и более рядов клеток.Они характерны для большинства лиственных пород. Ложноширокие сердцевинные лучи состоят из нескольких сближённых однорядных лучей (граб, ольха, лещина.). По высоте сердцевинные лучи у большинства пород состоят из нескольких рядов клеток. У некоторых пород (бук, липа, платан и др.) по высоте насчитывается более сотни рядов клеток. У большинства пород (дуб, ясень, береза, ольха и др.) на радиальном срезе все клетки луча несколько вытянуты вдоль его продольного направления. У некоторых пород встречаются неоднородные сердцевинные лучи. В них клетки верхних и нижних рядов слегка вытянуты вдоль оси ствола или имеют квадратную форму, а остальные клетки промежуточных рядов слегка вытянуты вдоль луча. Клетки сердцевинных лучей имеют многочисленные простые поры. Рассмотренные выше особенности микроскопического строения сердцевинных лучей являются важными диагностическими признаками для определения породы древесины.
Древесная паренхима на продольных срезах представлена отдельными вытянутыми вдоль ствола рядами паренхимных клеток.
С возрастом в стволе многолетних деревьев происходит ряд анатомических изменений. Покрывающая их перидерма заменяется более толстой и надежной коркой. Образование корки на стволах и старых скелетных ветвях у растений разных видов происходит в разном возрасте. Так как феллоген закладывается в коре все глубже и глубже, то вся первичная кора, а затем и более старые (периферийные) участки вторичной коры вынужденно отмирают и переходят в состав корки. В результате у старых стволов кора представлена относительно тонким слоем вторичной флоэмы. Ежегодно количество образуемой камбием работающей флоэмы примерно равно количеству уже не функционирующей флоэмы, которое переходит в состав корки. Таким образом, толщина луба, располагающегося между камбием и коркой, в течение всей дальнейшей жизни ствола остается почти неизменной. Нарастая изнутри, корка постепенно разрушается снаружи. Ее внешняя старая часть растрескивается под давлением растущего в толщину ствола и, распадаясь на фрагменты, постепенно опадает. Поэтому с возрастом толщина корки у ствола дерева увеличивается незначительно.
Постепенно прекращают проводить воду более старые, располагающиеся ближе к центру ствола участки древесины. У ряда лиственных пород (грецкий орех, дуб, робиния, или белая акация, шелковица) сосуды и трахеиды закупориваются тылами, у остальных (береза, вишня, клен и др.) — минеральными или органическими веществами (продуктами обмена веществ). Перемещению воды по трахеидам хвойных препятствует живица. Перекрывающие ток воды структуры и вещества не только представляют собой преграду на пути растущих гиф грибов- паразитов, но и защищают древесину от загнивания. Откладывающиеся во всех элементах старой древесины дубильные вещества (обладающие бактерицидными свойствами) часто окрашивают ее в темный цвет.
У ряда древесных пород (вяз, граб, дуб, ясень и др.) находящаяся в центральной части ствола более старая древесина в результате перечисленных изменений может хорошо отличаться от более молодой и еще функционирующей древесины более темной окраской и большей прочностью. Такую нефункционирующую древесину называют ядровой, или просто ядром. Остальную — более молодую и еще функционирующую древесину, которая окружает ядро, называют заболонью. Растения, образующие ядровую древесину, называют породами с твердой древесиной. Ядровая древесина начинает образовываться у растений в разное время: у дуба — в девять-десять лет, у сосны — в 25—30. У многих растений она очень красива. Ядровая древесина вишни — темно-красная, сливы — розоватая, дуба — коричневая, барбариса и бересклета — желтая.
Образование корки и формирование ядровой древесины имеют важное значение для дерева — они значительно продлевают его жизнь.
Растения, не образующие ядровой древесины, называют породами с мягкой древесиной (бук, ель, ива, липа, осина, тополь и др.). У этих растений более старая, переставшая проводить воду древесина внешне не отличается от функционирующей. Эту сухую древесину называют спелой. Обычно она мягкая, довольно легко режется и легко поддается разрушительной деятельности бактерий и грибов. Породы со спелой древесиной называют спелодревесными. В стволах таких деревьев легко образуются дупла. Недостаток одного из ценнейших растений, широко используемого при создании искусственных ландшафтов, — липы как раз и заключается в мягкой, легко разрушающейся под действием патогенных организмов древесине. Не подвергавшиеся своевременной грамотной формировке деревья становятся дуплистыми и начинают представлять опасность для посетителей парка или сквера.
11.Строение коры дерева. Возрастные изменения коры.
Древесная кора — комплекс высокоспециализированных клеток и тканей, располагающихся с внешней стороны от камбия и выполняющих защитную и проводящую функции. По проводящим элементам коры осуществляется транспорт питательных веществ, образующихся в листьях. Древесная кора защищает дерево от повреждения животными, дереворазрушающими насекомыми и организмами, вызывающими гниение.
Древесная кора также предохраняет камбий от потери влаги. По строению и составу древесная кора существенно отличается от древесины (ксилемы). Особая роль зеленых частей дерева — листвы и хвои, связанная с обеспечением жизненных процессов в растениях, в том числе древесных, также приводит к определенным особенностям их химического состава и строения.
В древесной коре, как и в древесине, сначала возникают первичные ткани, а затем при делении клеток вторичных меристем — камбия и пробкового камбия — образуются вторичные ткани, которые впоследствии отмирают. Наружная часть древесной коры — корка — состоит в основном из мертвых тканей и поэтому физиологически не активна.
В начале роста дерева из первичной верхушечной меристемы, наряду с первичной боковой меристемой – прокамбисм образуются первичные покровные ткани эпидермис и расположенная под ним первичная древесная кора, состоящая из слоев колленхимы и паренхимы. У молодых деревьев и побегов эпидермис состоит из одного ряда клеток эпидермы, покрытого снаружи гидрофобным воскоподобным веществом кутином. Колленхима состоит из клеток с утолщенными нелигнифицированными стенками и выполняет опорную (механическую) функцию. Из прокамбия в результате деления его клеток формируются первичная флоэма и первичная ксилема.
К концу первого вегетационного периода начинается вторичный рост. Из прокамбия образуется вторичная боковая меристема — камбий, а из него, в свою очередь, вторичная ксилема и флоэма. Под эпидермой появляется тонкий слой пробкового камбия (феллогена), в результате деления клеток которого образуется новая ткань перидерма. Эпидермис постепенно разрушается и в конце концов полностью заменяется перидермой, дающей начало внешнему покровному слою древесной коры. Перидерма состоит из трех слоев: пробкового камбия (феллогена); пробковой паренхимы (феллодермы); пробковой ткани (феллемы). Феллодерма образуется в результате деления клеток феллогена с внутренней стороны, афеллема с внешней стороны. Клетки феллодермы — паренхимные клетки, похожие на клетки лубяной паренхимы. Феллодерма развита меньше, чем феллема.
Процесс образования перидермы многообразен. У ряда древесных пород феллоген продолжает функционировать длительное время, обеспечивая равномерное нарастание слоя феллемы, что приводит к образованию толстого слоя пробки вместо типичной корки, как, например, у пробкового дуба, а также у дугласовой пихты, или к образованию гладкого эластичного наружного слоя древесной коры, как, например, у березы, осины, пихты. Стенки клеток пробки (феллемы) отличаются особым строением и составом. Они имеют три слоя. Наружный слой лигнифицирован, внутренний слой состоит практически из чистой целлюлозы, а средний слой содержит характерное для пробковой ткани вещество — суберин, причем слои суберина чередуются со слоями пробкового воска, что и обеспечивает гидрофобность пробки. В стенках клеток пробковой ткани березы содержится бетулин, придающий наружному слою коры березы — бересте — характерный белый цвет.
У большинства древесных пород, начиная с определенного возраста, слой пробковой ткани отмирает, а в глубине древесной коры закладываются новые слои перидермы. Во флоэме происходят изменения, связанные со старением и несколько напоминающие процесс образования ядровой древесины. Во внешней части флоэмы наблюдается так называемая облитерация — сплющивание ситовидных клеток или трубок и закупоривание их пористых пластинок, в результате чего первичная флоэма полностью отмирает.
Облитерированная вторичная флоэма при этом прерывается появляющимися слоями новой перидермы, имеющими неправильную форму. В этом процессе клетки феллогена образуются в результате деления живых паренхимных клеток флоэмы, возобновляющих меристематическую активность. Новый слой феллогена в свою очередь дает новые слои феллодермы и феллемы с последующим отмиранием клеток пробки и т.д. В результате такого процесса образуется сложный неоднородный комплекс тканей, состоящих преимущественно из мертвых клеток, внешняя основная часть корки (ритидом). Этот слой имеет характерный вид, изрезан трещинами. У различных видов сосны кора снаружи образует чешуйки. По мере роста деревьев в толщину кора снаружи постепенно отслаивается.
12.Корень, его зоны, первичное и вторичное строение.
Корень – осевой вегетативный, в типичном случае подземный орган сосудистых растений. Он эволюционировал позднее побега и устроен проще стебля. От побега корень отличается отсутствием листьев и их зачатков и тем, что его апекс (верхушка) прикрыт чехликом. Этот орган обладает радиальной симметрией и способностью неопределённо долгого нарастания в длину благодаря деятельности апикальной меристемы. Его главная функция – обеспечение почвенного питания растения. Кроме основной, корни решают и другие задачи:
- укрепление («заякоривание») растений в почве, что способствует росту побегов вверх и вынесению их к свету;
- синтезирование веществ – алкалоидов, аминокислот, гормонов и др., которые доставляются к другим органам растения;
- запасание веществ;
- вегетативное размножение;
- взаимодействиес корнями других растений, микроорганизмами, микоризными грибами.
Различные части корня выполняют неодинаковые функции и характеризуются определенными морфологическими особенностями. Эти части получили название зон корня:

– I зона деления – апикальная меристема корня (от 1 до 5 мм), в результате интенсивного деления ее клеток формируются все прочие зоны и ткани корня; от повреждений защищена корневым чехликом (1)– постоянно обновляющимся образованием на верхушке растущего корня – представлен особой паренхимной тканью, которая защищает апикальную меристему от трения о почвенные частицы и способствует продвижению корня благодаря выделению слизи; водные растения, как правило, корневого чехлика не имеют (вместо него формируется чехликоподобное образование – корневой кармашек);
– II зона роста, или растяжения – зона роста клеток (несколько мм), растягиваясь в продольном направлении, проталкивает корневое окончание вглубь почвы; клетки зоны характеризуются высоким тургором; в этой зоне начинается дифференциация клеток;
– III зона всасывания (от нескольких мм до нескольких см) ответственна за поглощение воды и минеральных веществ при помощи корневых волосков – выростов клеток эпиблемы (ризодермы) продолжительность жизни корневых волосков – 10–20 дней;
У одного растения ржи примерно 14 млрд. корневых волосков суммарной длиной более 10 000 км.
– IV зона проведения обладает хорошо развитой проводящей тканью, передает почвенные растворы в вышележащие отделы растения; корневые волоски вместе с клетками ризодермы погибают, и формируется покровная ткань, защищающая живые ткани корня; здесь появляются боковые корни (отсюда и второе название зоны – зона ветвления); составляет основную массу корня.
Резких границ между зонами корня нет.
Место перехода корня в стебель называется корневой шейкой.
Снаружи молодые корневые окончания покрыты эпиблемой (ризодермой) – однослойной первичной покровной тканью корня (образуется из дерматогена – наружный слой верхушечной меристемы кончика корня) (рис. 5.4.). В зоне поглощения клетки эпиблемы образуют корневые волоски, а в зоне проведения они довольно быстро слущиваются. Проникновение воды и солей в корневые волоски и далее в клетки коры и центрального цилиндра происходит путем осмоса, диффузии и активного переноса.
Под эпиблемой располагается первичная кора (дифференцируется из периблемы – периферийный отдел верхушечной меристемы, лежащий глубже дерматогена).
Наружные клетки первичной коры, лежащие непосредственно под эпиблемой, называются экзодермой.
Она бывает однослойной и многослойной (2 – 3 слоя). В зоне проведения после слущивания эпиблемы экзодерма оказывается снаружи, может опробковевать и выполняет функцию защитной покровной ткани.
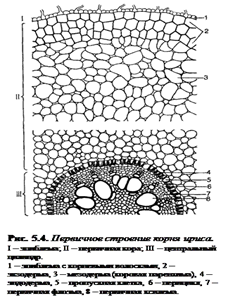
Основная масса первичной коры – мезодерма –образована паренхимными клетками: многослойная, рыхлая, с межклетниками. Через нее проходит радиальный (ближний) транспорт воды с минеральными веществами от эпиблемы к центральному цилиндру; здесь осуществляется активный синтез метаболитов и откладываются запасные питательные вещества.
Самым внутренним слоем коры является эндодерма, которая выполняет роль барьера – контролирует передвижение веществ из коры в осевой цилиндр и обратно. На ранних этапах развития эндодерма состоит из живых, тонкостенных клеток, расположенных плотно, без межклетников. Позднее ее клетки приобретают некоторые характерные структурные особенности: на радиальных стенках клеток эндодермы появляются особые утолщения (в результате суберинизации и одревеснения) – пояски Каспари, с помощью которых перекрывается передвижение растворов вдоль клеточных стенок (апопластный путь). В результате этого все вещества в центральный цилиндр и из него могут проникать только через живые протопласты клеток эндодермы (симпластный путь) и под их контролем (избирательная проницаемость, защита от проникновения болезнетворных микроорганизмов). У однодольных в клетках эндодермы могут происходить дальнейшие изменения: на внутренней поверхности первичных оболочек клеток откладывается суберин и далее вторичная целлюлозная оболочка, которая со временем одревесневает, таким образом перекрывается и второй путь передвижения веществ – симпластный (через цитоплазму клеток). На некоторых клетках этого не происходит, и они остаются только с поясками Каспари, это так называемые пропускные клетки – у однодольных только через них осуществляется физиологическая связь между первичной корой и осевым цилиндром. Пропускных клеток в эндодерме мало, а ее диаметр намного меньше, чем внешний диаметр корня в области эпиблемы, поэтому вокруг эндодермы образуется повышенное в сравнении с другими тканями количество воды. В результате этого вода проникает через пропускные клетки в сосуды ксилемы с определенным давлением, которое получило название корневого давления.
Центральный (осевой) цилиндр (формируется из плеромы – внутренней части апикальной меристемы) начинает дифференцироваться в зоне роста, вплотную к зоне деления. Наружный его слой – перицикл представляет собой однослойную образовательную ткань, состоящую из живых паренхимных клеток. В перицикле закладываются боковые корни, а также у некоторых растений возникают зачатки придаточных почек. У двудольных растений он участвует во вторичном утолщении корня, отчасти образуя камбий и феллоген.
Центральную часть цилиндра занимает сосудистоволокнистый пучок, состоящий из первичной ксилемы и первичной флоэмы (проводящие ткани образуются из прокамбия, который закладывается под перициклом). Элементы флоэмы и ксилемы закладываются по кругу, чередуясь друг с другом, и развиваются центростремительно (по направлению к центру корня), однако ксилема растет быстрее и занимает центр корня – сформировавшаяся структура проводящей ткани получила название радиального проводящего пучка.
Сердцевина
не типична для корня, но иногда (например, у кукурузы) заметна в центре в
виде небольшого участка механической ткани или тонкостенных клеток,
возникающих из прокамбия.
Такое первичное строение
корня сохраняется до конца жизни у хвощей, плаунов, папоротников и однодольных,
а у остальных – только в зоне всасывания.
Для голосеменных и двудольных растений характерны вторичные изменения в строении корня, обеспечивающие его рост в толщину. Вторичные изменения происходят за счет вторичных меристем – камбия и феллогена (рис. 5.7.).
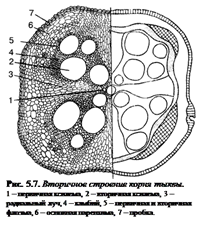
Вначале камбий появляется из клеток паренхимы центрального цилиндра (прокамбия) между сосудами первичной ксилемы и первичной флоэмы (с внутренней стороны). Участки камбия постепенно разрастаются, огибают флоэму и соприкасаются с перициклом. Клетки камбия прокамбиального происхождения делятся и дифференцируются: к центру откладывается вторичная ксилема, к наружи – вторичная флоэма. В местах, где сосуды первичной ксилемы соприкасаются с перициклом, клетки последнего также превращаются в камбий (перициклического происхождения), но он дифференцируется только в паренхиму, которая образует радиальные лучи напротив первичной ксилемы. Образуется камбиальное кольцо.
Вследствие быстрого нарастания изнутри вторичных тканей, обусловливающего сильное утолщение корня, первичная кора нередко разрывается.
Перицикл также становится меристематически активным: он делится и в центробежном направлении откладывает пробковый камбий – феллоген. Феллоген делится и кнаружи откладывает пробку (феллему) – вторичную покровную ткань корня. Пробка изолирует первичную кору от проводящих тканей, она теряет связь с живыми клетками центрального цилиндра, отмирает и слущивается. Внутрь феллоген откладывает феллодерму, которая затем превращается в паренхимную ткань и образует вместе с перициклом вторичную кору. Снаружи корни двудольных растений, имеющие вторичное строение, покрыты перидермой. Корка образуется редко, лишь на старых корнях деревьев.
Многолетние корни древесных растений в результате длительной камбиальной активности нередко сильно утолщаются. Вторичная ксилема у таких корней сливается в сплошной цилиндр, окруженный снаружи кольцом камбия и сплошным кольцом вторичной флоэмы.
Таким образом, этапы перехода корня от первичного строения ко вторичному следующие:
1. Появление камбия между лучами ксилемы и флоэмы и образование камбиального кольца;
2. Образование феллогена перициклом, возникновение вторичной коры;
3. Сбрасывание первичной коры;
4. Смена радиального расположения тканей ксилемы и флоэмы коллатеральным.
13.Анатомическое строение плоского и иглового листа, листьев светового и теневого типа.
Типичный лист представляет собой боковой вырост стебля и состоит из черешка и листовой пластинки. В листе имеется верхняя брюшная и нижняя спинная сторона (лат. dorsum ≈ спина, venter ≈ брюхо).
По своей физиологической роли лист - орган фотосинтеза, газообмена и транспирации. В тканях листа совершается превращение неорганических веществ (СО2 и Н2О) в органические. Продукты фотосинтеза обычно не накапливаются в листе, а перетекают в другие органы растения.
Основной, типичной категорией листьев являются зеленые, ассимилирующие (питающие) листья, которые называются трофофиллам. Листья являются пластичными, изменчивыми, в эволюционном отношении органами. В листе хорошо представлены следующие ткани:
|
- питательная или ассимиляционная; |
|
|
- покровная, регулирующая испарение воды и газообмен; |
|
|
- проводящие такни - обеспечивают подведение почвенных растворов и отток продуктов ассимиляции; |
|
|
- механические ткани, придающие листу прочность. |
Кроме этих четырех тканей, в листе могут встречаться группы клеток или отдельные клетки идиобласты: склереиды, млечники, места отложения отбросов минеральных солей и других специфических веществ.
Листья возникают экзогенно (поверхностно), в виде бугорков вблизи от точки роста.
Лист покрыт со всех сторон эпидермой (кожицей). В обычных, горизонтально ориентированных листьях верхний и нижний эпидермис различаются.
В целом, кожица состоит из тесно сомкнутых паренхимных клеток с извилистыми очертаниями. Клетки нижнего эпидермиса обычно более мелкие и более извилистые.
В верхнем эпидермисе в типичном случае сильнее развиты кутикулярные слои, однако опушение приурочено главным образом к нижней стороне листа. Здесь же, в нижнем эпидермисе обычно располагаются и устьица.
Листья хвойных игольчатые (сосна, ель, пихта, лиственница) или чешуевидные (туя, кипарис), как правило, вечнозелёные (искл. Лиственница – вторичное приспособление к очень холодному климату), приспособлены к экономной транспирации воды и к перенесению засухи, в том числе и зимней, когда при низких температурах корни не могут поглощать воду.
1) Наружная поверхность хвоинок очень мала (мала площадь испарения).
2) Эпидерма состоит из толстостенных клеток с мощной кутикулой (защита от испарения).
3) Погружённые устьица. Замыкающие клетки частично одревесневают, а канал заполняется смолами или воском (резкое снижение транспирации).
4) Под эпидермой кольцом расположена особая ткань гиподерма, состоящая из одревесневших волокон, что снижает испарение и повышает механическую прочность.
5) Главное отличие от покрытосеменных: нет дифференциации на столбчатый и губчатый мезофилл, все клетки однородные, образуют складчатый мезофилл. Это адаптивная компенсация малой наружной поверхности. У клеток мезофилла оболочка образует внутренние складки, что обеспечивает резкое возрастание постенного слоя цитоплазмы и внутренней поверхности.
В клетках за счёт увеличения их внутренней поверхности возрастает число хлоропластов, и при малой внешней поверхности хвоинки процессы фотосинтеза идут также интенсивно, как и в обычных листьях цветковых растений.
6) Двойной сосудисто-волокнистый пучок окружён эндодермой, регулирующей транспорт веществ. При вхождении в стебель двойной пучок сливается в один, образуя один листовой след.
7) Проводящие пучки окружены трансфуззионной тканью, которая состоит: а) из лучевых трахеид (транспорт воды), б) клеток живой паренхимы (транспорт органических веществ-ассимилятов).
8) Имеются смоляные ходы, расположенные между клетками мезофилла.
У растений, обитающих в условиях хорошей освещенности, листья содержат несколько слоев столбчатых клеток. Хорошо развита в них и губчатая ткань. Такие листья называют световыми. У теневыносливых растений листья имеют по одному слою мелких столбчатых клеток, и губчатая ткань у них менее развита. Такие листья называют теневыми. Таким образом, листовые пластинки у световых листьев толще, чем у теневых, однако в теневых листьях хлоропласты крупнее и содержат больше хлорофилла. Поэтому они и имеют темно-зеленую окраску. Световые листья — светло-зеленые, так как в них содержится меньше хлорофилла. Световые и теневые листья отличаются размерами. В лесах растения имеют крупные листья, которые улавливают больше света.
 (zip - application/zip)
(zip - application/zip)